Процесс реализации генетической информации. Этапы реализации генетической информации
Сложившиеся представления о внутриклеточном переносе генетической информации по схеме ДНК->РНК->белок, предложенной Ф. Криком, принято называть «центральной догмой » молекулярной биологии. Наряду с этим (наиболее распространенным) направлением переноса, который иногда обозначают как общий перенос, известна и другая форма реализации генетической информации (специализированный перенос), обнаруженная при инфицировании клетки РНК-co держащими вирусами. В этом случае наблюдается процесс, получивший название обратной транскрипции, при котором первичный генетический материал (вирусная РНК), проникший в клетку хозяина, служит матрицей для синтеза комплементарной ДНК с помощью фермента обратной транскриптазы, кодируемой вирусным геномом. В дальнейшем возможна реализация информации синтезированной вирусной ДНК в обычном направлении. Следовательно, специализированный перенос генетической информации осуществляется по схеме РНК-»ДНК-»РНК-»белок.
Транскрипция является первым этапом общего переноса генетической информации и представляет собой процесс биосинтеза молекул РНК на матрице ДНК. Принципиальный смысл этого процесса состоит в том, что информация структурного гена (либо нескольких расположенных рядом генов), записанная в форме нуклеотидной последовательности матричной нити ДНК (5’, переписывается (транскрибируется) в нуклеотидную последовательность молекулы РНК, синтезируемой в направлении 5’->3’ на основе комплементарного соответствия дезоксирибонуклеотидов цепи ДНК рибонуклеотидам РНК (А - У, Г - Ц, Т - А, Ц - Г). Вторая цепь ДНК, комплементарная матричной, называется кодирующей («-»-цепь).
В качестве продуктов транскрипции (транскриптов) можно рассматривать все типы клеточной РНК. Единица транскрипции получила название «транскриптон». На рисунке 1.4 представлена структура прокариотического транскриптона.
Рис. 1.4.
Процесс транскрипции катализируется РНК-полимеразой, представляющей собой сложный белок, состоящий из нескольких субъединиц и способный выполнять несколько функций.
Транскрипцию принято подразделять на три основных стадии: инициацию (начало синтеза РНК), элонгацию (удлинение полинуклеотидной цепочки) и терминацию (окончание процесса). Рассмотрим данный процесс на примере прокариотической клетки.
Инициация транскрипции осуществляется РНК-полимеразой в состоянии холофермента, т.е. в присутствии всех субъединиц (двух а, формирующих каркас РНК-полимеразы; р, катализирующей полимеризацию РНК; Р’, обеспечивающей неспецифическое связывание с ДНК; со, участвующей в сборке фермента и защищающей его от разрушения; о, распознающей промотор и связывающейся с промотором). Фермент связывается с участком ДНК, называемым промотором (рис. 1.5) и расположенным перед стартовой точкой, с которой начинается синтез РНК. Промоторы разных структурных генов могут быть идентичными либо содержат отличающиеся друг от друга последовательности нуклеотидов, что, вероятно, определяет эффективность транскрибирования отдельных генов и возможности регуляции самого процесса транскрипции. Промоторы большинства генов прокариот имеют в своем составе универсальную последовательность 5’-ТАТААТ-3’ (блок Прибнова), которая располагается перед стартовой точкой на расстоянии порядка десяти нуклеотидов и распознается РНК-полимеразой. Другая относительно часто встречающаяся узнаваемая последовательность этих организмов (5’-ТТГАЦА-3’) обычно обнаруживается на расстоянии примерно 35 нуклеотидов от стартовой точки. Специфическое прочное связывание РНК-полимеразы с тем или иным узнаваемым ею участком промоторной области позволяет ей начать процесс расплетания молекулы ДНК вплоть до стартовой точки, с которой она начинает осуществлять полимеризацию рибонуклеотидов с использованием в качестве матрицы одно- нитевого 3’-5’-фрагмента ДНК. После синтеза короткого (длиной до десяти нуклеотидов) фрагмента РНК, G-субъединица отсоединяется, и РНК-полимераза переходит в состояние кор-фермента.

Рис. 1.5.
На этапе элонгации кор-фермент продвигается по ДНК-матрице, расплетая ее и наращивая цепь РНК в направлении 5’->3’. Вслед за продвижением РНК-полимеразы восстанавливается исходная вторичная структура ДНК. Процесс продолжается вплоть до достижения РНК-полимеразой области терминатора. Последний представляет собой нуклеотидную последовательность ДНК, на которой оканчивается синтез транскрипта, и он отсоединяется от матрицы. Существуют два основных способа терминации. При р-независимой терминации на синтезируемой РНК формируется шпилька, препятствующая дальнейшей работе РНК-полимеразы, и транскрипция прекращается, p-зависимая терминация осуществляется с участием р-белка, который присоединяется к определенным участкам синтезируемой РНК и с затратой энергии АТФ способствует диссоциации гибрида РНК с матричной нитью ДНК. В большинстве случаев терминатор находится в конце структурного гена, обеспечивая синтез одной моногенной молекулы мРНК. Вместе с тем у прокариот возможен синтез полигенной молекулы мРНК, которая кодирует синтез не одного, а двух и большего числа полипептидных цепочек. В этом случае происходит непрерывное транскрибирование нескольких расположенных рядом друг с другом структурных генов, имеющих один общий терминатор. Однако полигенная мРНК может содержать в своем составе нетранслируемые межгенные области (спейсеры), разделяющие кодирующие участки для отдельных полипептидов, что, вероятно, обеспечивает последующее разделение и самих синтезируемых полипептидов.
В отличие от прокариот, в клетках которых имеется РНК-поли- мераза лишь одного типа, обеспечивающая синтез разных молекул РНК, у эукариот обнаружены ядерные РНК-полимеразы трех типов (I, II, III), а также РНК-полимеразы клеточных органелл, содержащих ДНК (митохондрий, пластид). РНК-полимераза I находится в ядрышке и участвует в синтезе большинства молекул рРНК (5,8S, 18S, 28S), РНК-полимераза II обеспечивает синтез мРНК, мяРНК и микроРНК, а РНК-полимераза III осуществляет синтез тРНК и 5S рРНК.
Разные типы РНК-полимераз инициируют транскрипцию с разных промоторов. Так, промотор для РНК-полимеразы II (рис. 1.6) содержит универсальные последовательности ТАТА (блок Хогнесса), ЦААТ и состоящие из повторяющихся нуклеотидов Г и Ц (ГЦ-моти- вы). При этом та или иная промоторная область может включать либо одну из указанных последовательностей, либо комбинацию двух или трех таких последовательностей. Также для инициации транскрипции эукариотические РНК-полимеразы нуждаются в белках - факторах транскрипции.

Рис. 1.6.
Поскольку структурные гены эукариот имеют прерывистое (мозаичное) строение, то их транскрипция имеет специфические особенности, отличающие ее от транскрипции у прокариот. На рисунке 1.7 представлена структура эукариотического транскриптона. В случае эукариотического гена, кодирующего синтез полипептида, этот процесс начинается с транскрибирования всей нуклеотидной последовательности, содержащей как экзонные, так и интронные участки ДНК. Образовавшаяся при этом молекула РНК, отражающая структуру всего мозаичного гена, которую называют гетерогенной ядерной РНК (гяРНК) либо проматричной РНК (про-мРНК), претерпевает затем процесс созревания (процессинг мРНК).

Рис. 1.7.
Процессинг мРНК у эукариот включает три этапа: кэпирование, полиаденилирование и сплайсинг. Модификация 5’-конца, называемая копированием, представляет собой присоединение к 5’-концу транскрипта гуанозинтрифосфата (ГТФ) необычной 5’-5’- связью. Реакция катализируется ферментом гуанилилтрансферазой. Затем происходит метилирование присоединенного гуанина и первых нуклеотидов транскрипта. Функциями «кэпа» (от англ, cap - колпачок, шапочка), вероятно, являются защита 5’-конца мРНК от ферментативной деградации, взаимодействие с рибосомой при инициации трансляции и транспорт мРНК из ядра. Модификация З’-конца (по- лиаденилирование) - это присоединение к З’-концу РНК-транскрип- та от 100 до 300 остатков адениловой кислоты. Процесс катализируется ферментом polyA-полимеразой. Для действия фермента, осуществляющего полиаденилирование, не нужна матрица, но требуется присутствие на З’-конце мРНК сигнальной последовательности ААУААА. Предполагается, что полиадениловый «хвост» обеспечивает транспорт зрелой мРНК к рибосоме, защищая ее от ферментативного разрушения, но сам постепенно разрушается ферментами цитоплазмы, отщепляющими один за другим концевые нуклеотиды. Третий этап процессинга - сплайсинг состоит в ферментативном разрезании первичного транскрипта с последующим удалением его интронных участков и воссоединением экзонных участков, формирующих непрерывную кодирующую последовательность зрелой мРНК, которая в дальнейшем участвует в трансляции генетической информации. В сплайсинге принимают участие короткие молекулы мяРНК, состоящие примерно из 100 нуклеотидов, которые представляют собой последовательности, являющиеся комплементарными последовательностям на концах интронных участков гяРНК. Спаривание комплементарных нуклеотидов мяРНК и первичного транскрипта способствует сворачиванию в петлю интронных участков и сближению соответствующих экзонных участков гяРНК, что, в свою очередь, делает их доступными разрезающему действию ферментов (нуклеаз). Следовательно, молекулы мяРНК обеспечивают правильность вырезания нитронов из гяРНК.
Следует отметить, что у эукариот процессингу подвергается большинство типов РНК, в то время как у прокариот мРНК процессингу не подвергается, и трансляция синтезируемой молекулы мРНК может начаться до завершения транскрипции.
Трансляция как очередной этап реализации генетической информации заключается в синтезе полипептида на рибосоме, при котором в качестве матрицы используется молекула мРНК (считывание информации в направлении 5’ -> 3’). В клетках прокариот генетический материал (ДНК) находится в цитоплазме, что определяет сопряженность процессов транскрипции и трансляции. Иными словами, образовавшийся лидирующий 5’-конец молекулы мРНК, синтез которой еще не завершен, уже способен вступать в контакт с рибосомой, инициируя синтез полипептида, т.е. транскрипция и трансляция идут одновременно. Что касается эукариот, то процессы транскрипции и трансляции разделены в пространстве и во времени в связи с процессингом молекул РНК и необходимостью их последующей транспортировки из ядра в цитоплазму, где будет осуществляться синтез полипептида.
Как и в случае транскрипции, процесс трансляции можно условно подразделить на три основных стадии: инициацию, элонгацию и терминацию.
Как известно, отдельная рибосома представляет собой клеточную органеллу, состоящую из молекул рРНК и белков (рис. 1.8). В составе рибосомы имеются две структурные субъединицы (большая и малая), которые можно дифференцировать на основании их способности по-разному осаждаться при ультрацентрифугировании препаратов очищенных рибосом из разрушенных клеток, т.е. по коэффициенту седиментации (величине S). При определенных условиях в клетке может происходить разделение (диссоциация) этих двух субъединиц либо их объединение (ассоциация).

Рис. 1.8.
Рибосомы прокариот состоят из большой и малой субъединиц с величинами 50S и 30S соответственно, тогда как у эукариот эти субъединицы крупнее (60S и 40S). Поскольку процесс трансляции более детально был исследован у бактерий, то и здесь мы его рассмотрим на примере прокариот. Как видно из рис. 1.8, в рибосоме содержатся несколько активных центров: A-участок (аминоацильный), P-участок (пептидильный), Е-участок (для выхода пустой тРНК) и участок связывания мРНК.
В процесс трансляции вовлечены также молекулы тРНК, функции которых состоят в участии в транспорте аминокислот из цитозоля к рибосомам и в распознавании кодона мРНК. Молекула тРНК, имеющая вторичную структуру в форме «клеверного листа», содержит в своем составе тройку нуклеотидов (антикодон), которая обеспечивает ее комплементарное соединение с соответствующим кодоном молекулы мРНК, и акцепторный участок (на З’-конце молекулы), к которому присоединяется определенная аминокислота (см. рис. 1.3). Каждая аминокислота, участвующая в процессе трансляции, прежде чем переместиться к рибосоме, должна присоединиться к определенной тРНК с помощью соответствующего варианта фермента аминоацил-тРНК-синтетазы с использованием энергии молекул АТФ. Образование комплекса аминоацил-тРНК проходит в два этапа.
- 1. Активация аминокислоты: Аминокислота + АТФ -> аминоа- цил-АМФ + РР.
- 2. Присоединение аминокислоты к тРНК: Аминоацил-АМФ + + тРНК -> аминоацил-тРНК + АМФ.
Инициация трансляции у прокариот сопровождается диссоциацией рибосомы на две субъединицы. Затем 5-8 нуклеотидная последовательность, расположенная на 5’-конце молекулы мРНК (последовательность Шайна - Далъгарно) связывается с определенной областью малой субъединицы рибосомы таким образом, что в P-участке оказывается стартовый (инициирующий) кодон АУГ этой молекулы. Функциональная особенность такого P-участка во время инициации состоит в том, что он может быть занят только инициирующей аминоацил-тРНК с антикодоном УАЦ, которая у эукариот несет аминокислоту метионин, а у бактерий - формилметионин. Поскольку синтез полипептида всегда начинается с N-конца и идет в направлении к С-концу, то все белковые молекулы, синтезируемые в клетках прокариот, должны начинаться с N-формилметионина, а у эукариот - с N-метионина. Однако в дальнейшем эти аминокислоты, как правило, ферментативно отщепляются во время процессинга белковой молекулы. После образования инициирующего комплекса в «недостроенном» P-участке становится возможным воссоединение малой и большой субъединиц рибосомы, что приводит к «достраиванию» Р-участка и А-участка.
Процесс элонгации начинается с доставки следующей аминоацил-тРНК в A-участок рибосомы и присоединения на основе принципа комплементарности ее антикодона к соответствующему кодону мРНК, находящемуся в этом участке. Затем образуется пептидная связь между инициирующей (первой в цепочке) и последующей (второй) аминокислотами, после чего происходит перемещение рибосомы на один кодон мРНК в направлении 5’ -» 3’, что сопровождается отсоединением инициирующей тРНК от матрицы (мРНК) и от инициирующей аминокислоты и выходом ее в цитоплазму через Е-участок.
При этом вторая по счету аминоацил-тРНК передвигается из А-участ- ка в P-участок, а освободившийся A-участок занимается следующей (третьей по счету) аминоацил-тРНК. Процесс последовательного передвижения рибосомы «триплетными шагами» по нити мРНК повторяется, сопровождаясь освобождением тРНК, поступающих в Р-участок, и наращиванием аминокислотной последовательности синтезируемого полипептида.
И инициация, и элонгация трансляции осуществляются с участием вспомогательных белковых факторов. На сегодняшний день у прокариот описано по три таких фактора для каждого из этапов синтеза белка.
Терминация трансляции связана с вхождением одного из трех известных стоп-кодонов мРНК (УАА, УАГ, УГА) в A-участок рибосомы. Поскольку эти кодоны не несут информации о какой-либо аминокислоте, но узнаются соответствующими факторами терминации, процесс синтеза полипептида прекращается, и он отсоединяется от матрицы (мРНК).
После выхода из функционирующей рибосомы свободный 5’-ко- нец мРНК может вступать в контакт со следующей рибосомой, инициируя синтез еще одного (идентичного) полипептида. Следовательно, рассмотренный рибосомный цикл последовательно повторяется с участием нескольких рибосом, в результате чего формируется структура, называемая полисомой и представляющая собой несколько рибосом, одновременно транслирующих одну молекулу мРНК.
Механизм синтеза полипептида в эукариотической клетке принципиально схож с таковым у прокариот. Однако отличаются вовлеченные в процесс белковые факторы.
Посттрансляционная модификация полипептида представляет собой завершающий этап реализации генетической информации в клетке, приводящий к превращению синтезированного полипептида в функционально активную молекулу белка. При этом первичный полипептид может претерпевать процессинг, состоящий в ферментативном удалении инициирующих аминокислот, отщеплении других (ненужных) аминокислотных остатков и в химической модификации отдельных аминокислот. Затем происходит процесс сворачивания линейной структуры полипептида за счет образования дополнительных связей между отдельными аминокислотами и формирование вторичной структуры белковой молекулы. На этой основе формируется еще более сложная третичная структура молекулы.
В случае белковых молекул, состоящих более чем из одного полипептида, происходит образование комплексной четвертичной струк- зв туры, в которой объединяются третичные структуры отдельных полипептидов. В качестве примера можно привести молекулу гемоглобина человека, состоящую из двух а-цепочек и двух (3-цепочек, которые формируют стабильную тетрамерную структуру. Каждая из глобино- вых цепочек содержит также молекулу гема, который в комплексе с железом способен связывать молекулы кислорода, обеспечивая их транспортировку эритроцитами крови.
ЗАДАНИЯ И ВОПРОСЫ ДЛЯ САМОСТОЯТЕЛЬНОЙ РАБОТЫ
1. Фрагмент кодирующей цепи ДНК имеет следующую нуклеотидную последовательность: 5’-ГАТТЦТГАЦТЦАТТГЦАГ-3’
Определите ориентацию и нуклеотидную последовательность мРНК, синтезируемой на указанном фрагменте ДНК, и аминокислотную последовательность кодируемого ею полипептида.
- 2. Можно ли однозначно определить нуклеотидную последовательность мРНК и комплементарной ей нити ДНК, если известна аминокислотная последовательность кодируемого ими полипептида? Дайте обоснование своего ответа.
- 3. Запишите все варианты фрагментов мРНК, которые могут кодировать следующий фрагмент полипептида: Фен - Мет - Цис.
- 4. Какие аминокислоты могут транспортировать к рибосомам тРНК с антикодонами: АУГ, ААА, ГУЦ, ГЦУ, ЦГА, ЦУЦ, УАА, УУЦ?
- 5. Как можно объяснить то обстоятельство, что размеры нуклеотидной последовательности структурного гена (3-глобина (1380 пар нуклеотидов) значительно превышают величину, необходимую для кодирования соответствующего полипептида, состоящего из 146 аминокислотных остатков?
Вопрос 1. Вспомните полное определение понятия «жизнь».
В середине XIX в. Фридрих Энгельс писал: «Жизнь есть способ существования белковых тел, существенным моментом которого является постоянный обмен веществ с окружающей их внешней природой, причем с прекращением этого обмена веществ прекращается и жизнь, что приводит к разложению белка». На современном уровне знаний это классическое определение жизни дополнено представлением об исключительной значимости нуклеиновых кислот - молекул, которые содержат генетическую информацию, позволяющую организмам самовозобновляться и самовоспроизводиться (размножаться).
Приведем одно из современных определений,. данное советским учёным-биологом М.В.Волькенштейном 1965 г.), «Живые тела, существующие на Земле, представляют собой открытые, саморегулирующиеся и самовоспроизводящиеся системы, построенные из биополимеров - белков и нуклеиновых кислот». При этом понятие «открытая система» подразумевает отмеченный еще Ф. Энгельсом обмен веществами и энергией с окружающей средой (питание, дыхание, выделение); понятие «саморегуляция» - способность к поддержанию постоянства химического состава, структуры и свойств. Важным условием успешной саморегуляции является раздражимость - способность организма реагировать на информацию, поступающую из внешнего мира.
Вопрос 2. Назовите основные свойства генетического кода и поясните их значение.
Генетический код – это последовательность нуклеотидов в ДНК, контролирующая последовательность аминокислот в белковой молекуле.
Свойства кода
1. Триплетность. Одну аминокислоту кодирует три нуклеотида, которые называют триплетом или кодоном.
2. Вырожденность или избыточность. Каждая аминокислота зашифрована более чем одним кодоном. Для кодирования 20 аминокислот (в основном столько входит в состав белка) используется 61 комбинация нуклеотидов (4 3 = 64). Три кодона: УАА, УАГ, УГА - называют триплетами терминации, т.е. они несут информацию о прекращении синтеза белка.
3. Универсальность. У всех организмов на Земле одни и те же триплеты кодируют одинаковые аминокислоты.
4. Однозначность. Каждый триплет кодирует только одну аминокислоту.
5. Колинеарность или линейность. Нуклеотиды в ДНК и и-РНК располагаются линейно и так же линейно будут расположены аминокислоты в белковой молекуле.
6. Неперекрываемость. Информация считывается триплетами, т.е. каждый нуклеотид входит в состав только одного кодона.
7. Полярность. Существуют триплеты, определяющие начало и конец отдельных генов. т.д.
Вопрос 3. Какова сущность процесса передачи наследственной информации из поколения в поколение и из ядра в цитоплазму, к месту синтеза белка?
При передаче наследственной информации из поколения в поколение молекулы ДНК удваиваются в процессе дупликации. Каждая дочерняя клетка получает одну из двух идентичных молекул ДНК. При бесполом размножении генотип дочернего организма идентичен материнскому. При половом размножении организм потомка получает собственный диплоидный набор хромосом, собранный из гаплоидного материнского и гаплоидного отцовского наборов.
При передаче наследственной информации из ядра в цитоплазму ключевым процессом является транскрипция - синтез РНК на ДНК. Синтезированная молекула иРНК является комплементарной копией определенного фрагмента ДНК - гена и содержит информацию о строении определенного белка. Такая молекула иРНК является посредником между хранилищем генетической информации - ядром и цитоплазмой с рибосомами, где создаются белки. Рибосомы используют иРНК как матрицу («инструкцию») для синтеза белка в процессе трансляции.
Вопрос 4. Где синтезируются рибонуклеиновые кислоты?
Рибонуклеиновые кислоты синтезируются в ядре. Образование рРНК и сборка субъединиц рибосом происходят в особых участках ядра - ядрышках. Небольшое количество РНК синтезируется в митохондриях и пластидах, где имеется собственная ДНК и собственные рибосомы.
Вопрос 5. Расскажите, где происходит синтез белка и как он осуществляется.
Синтез белка происходит в цитоплазме и осуществляется с помощью специализированных органоидов - рибосом. Молекула иРНК соединяется с рибосомой тем концом, с которого должен начаться синтез белка. Аминокислоты, необходимые для синтеза белковой цепи, доставляются молекулами транспорт¬ных РНК (тРНК). Каждая тРНК может переносить только одну из 20 аминокислот (например, только цистеин). Какую конкретно аминокислоту переносит тРНК, определяет триплет нуклеотидов, расположенный на верхушке центральной петли тРНК, - антикодон. Если антикодон окажется комплементарен триплету нуклеотидов иРНК, находящемуся в данный момент в контакте с рибосомой, про¬изойдет временное связывание тРНК с иРНК, и аминокислота будет включена в белковую цепь.BR> На следующем этапе освободившаяся тРНК уйдет в цитоплазму, а рибосома сделает «шаг» и сдвинется к следующему триплету иРНК. Затем к этому триплету подойдет тРНК с соответствующим антикодоном и доставит очередную аминокислоту, которая будет присоединена к растущему белку.
Таким образом, включение аминокислот в белковую цепь происходит строго в соответствии с последовательностью расположения триплетов цепи иРНК.
Всю последовательность процессов, происходящих при синтезе белковых молекул, можно объединить в три этапа:
I Транскрипция
- (лат. transcriptio
-переписывание)- это переписывание информации по принципу комплементарности с ДНК на и-РНК.
II Процессинг
- (лат. processing
- обработка) – это созревание и-РНК.
В результате процессинга образуется короткая зрелая и-РНК или еще ее называют матричная (м-РНК). Эти два этапа идут в ядре. Через ядерные поры зрелая короткая и-РНК выходит в цитоплазму.
III Трансляция
- (лат. translatio
– перевод) - это синтез на рибосомах полипептидных цепей. На и-РНК может объединиться несколько рибосом и такая структура называется полирибосома или полисома. Аминокислоты, из которых синтезируются белки, доставляются к рибосомам с помощью т-РНК. К основанию т-РНК присоединяется аминокислота, которая кодируется антикодоном.
Специфичность белка определяется порядком, количеством и разнообразием аминокислот, входящих в его состав.
Сначала, установите последовательность этапов биосинтеза белка, начиная с транскрипции. Всю последовательность процессов, происходящих при синтезе белковых молекул, можно объединить в 2 этапа:
Транскрипция.
Трансляция.
Структурными единицами наследственной информации являются гены – участки молекулы ДНК, кодирующие синтез определенного белка. По химической организации материал наследственности и изменчивости про- и эукариот принципиально не отличается. Генетический материал в них представлен в молекуле ДНК, общим является также принцип записи наследственной информации и генетический код. Одни и те же аминокислоты у про — и эукариот шифруются одинаковыми кодонами.
Геном современных прокариотических клеток характеризуется относительно небольшими размерами, ДНК кишечной палочки имеет вид кольца, длиной около 1 мм. Она содержит 4 х 10 6 пар нуклеотидов, образующих около 4000 генов. В 1961 г. Ф. Жакоб и Ж. Моно открыли цистронную, или непрерывную организацию генов прокариот, которые полностью состоят из кодирующих нуклеотидных последовательностей, и они целиком реализуются в ходе синтеза белков. Наследственный материал молекулы ДНК прокариот располагается непосредственно в цитоплазме клетки, где также находятся необходимые для экспрессии генов тРНК и ферменты.Экспрессия- это функциональная активность генов, или выражение генов. Поэтому синтезированная с ДНК иРНК способна сразу выполнять функцию матрицы в процессе трансляции синтеза белка.
Геном эукариот содержит значительно больше наследственного материала. У человека общая длина ДНК в диплоидном наборе хромосом составляет около 174 см. Она содержит 3 х 10 9 пар нуклеотидов и включает до 100000 генов. В 1977 г. была обнаружена прерывистость в строении большинства генов эукариот, получивший название «мозаичный» ген. Для него характерны кодирующие нуклеотидные последовательности экзонные и интронные участки. Для синтеза белка используется только информация экзонов. Количество интронов варьирует в разных генах. Установлено,что ген овальбумина кур включает 7 интронов, а ген проколлагена млекопитающих – 50. Функции молчащей ДНК – интронов окончательно не выяснены. Предполагают, что они обеспечивают: 1) структурную организацию хроматина; 2) некоторые из них, очевидно, участвуют в регуляции экспрессии генов; 3) интроны можно считать запасом информации для изменчивости; 4) они могут играть защитную роль, принимая на себя действие мутагенов.
Транскрипция
Процесс переписывания информации в ядре клетки с участка молекулы ДНК на молекулу мРНК (иРНК) называется транскрипция (лат. Transcriptio – переписывание). Синтезируется первичный продукт гена- мРНК. Это первый этап белкового синтеза. На соответствующем участке ДНК фермент РНК–полимераза узнает знак начала транскрипции – промотр. Стартовой точкой считается первый нуклеотид ДНК, который включается ферментом в РНК-транскрипт. Как правило, кодирующие участки начинаются кодоном АУГ, иногда у бактерий используется ГУГ. Когда РНК-полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и копируется одна из цепей по принципу комплементарности. Синтезируется мРНК, скорость сборки её достигает 50 нуклеотидов в секунду. По мере движения РНК–полимеразы, растёт цепь мРНК, и когда фермент достигнет конца копирующего участка – терминатора , мРНК отходит от матрицы. Двойная спираль ДНК позади фермента восстанавливается.
Транскипция прокариот осуществляется в цитоплазме. В связи с тем, что ДНК целиком состоит из кодирующих нуклеотидных последовательностей, поэтому синтезированная мРНК сразу выполняет функцию матрицы для трансляции (см. выше).
Транскрипция мРНК у эукариот происходит в ядре. Она начинается синтезом больших по размерам молекул — предшественников (про-мРНК), называемых незрелой, или ядерной РНК.Первичный продукт гена- про-мРНК является точной копией транскрибированного участка ДНК, включает экзоны и интроны. Процесс формирования зрелых молекул РНК из предшественников называется процессингом . Созревание мРНК происходит путём сплайсинга – это вырезания ферментами рестриктаз интронов и соединение участков с транскрибируемыми последовательностями экзонов ферментами лигаз. (Рис.).Зрелая мРНК значительно короче молекул-предшественников про – мРНК, размеры интронов в них варьирует от 100 до 1000 нуклеотидов и более. На долю интронов приходится около 80% всей незрелой мРНК.
В настоящее время доказана возможность альтернативного сплайсинга, при котором из одного первичного транскрипта могут удалятся в разных его участках нуклеотидные последовательности и будут образовываться несколько зрелых мРНК. Данный вид сплайсинга характерен в системе генов иммуноглобулинов у млекопитающих, что даёт возможность формировать на основе одного транскрипта мРНК разные виды антител.
По завершению процессинга зрелая мРНК проходит отбор перед выходом из ядра. Установлено, что в цитоплазму попадает всего 5% зрелой мРНК, а остальная часть расщепляется в ядре.
Трансляция
Трансляция (лат. Translatio — передача, перенесение) — перевод информации, заключенной в последовательности нуклеотидов молекулы мРНК,в последовательность аминокислот полипептидной цепи (Рис. 10). Это второй этап белкового синтеза. Перенос зрелой мРНК через поры ядерной оболочки производят специальные белки, которые образуют комплекс с молекулой РНК. Кроме транспорта мРНК, эти белки защищают мРНК от повреждающего действия цитоплазматических ферментов. В процессе трансляции центральная роль принадлежит тРНК, они обеспечивают точное соответствие аминокислоты коду триплета мРНК. Процесс трансляции- декодирования происходит в рибосомах и осуществляется в направлении от 5 к 3 , Комплекс мРНК и рибосом называется полисомой.
В ходе трансляции можно выделить три фазы: инициацию, элонгацию и терминацию.
Инициация.
На этом этапе происходит сборка всего комплекса, участвующего в синтезе молекулы белка. Происходит объединение двух субъединиц рибосом на определённом участке мРНК, присоединение к ней первой аминоацил – тРНК и этим задаётся рамка считывания информации. В молекуле любой м-РНК есть участок, комплементарный р-РНК малой субединицы рибосомы и специфически ею управляемый. Рядом с ним находится инициирующий стартовый кодон АУГ, который кодирует аминокислоту метионин.Фаза инициации завершается образованием комплекса:рибосома, -мРНК- инициирующая аминоацил-тРНК.
Элонгация
— она включает все реакции от момента образования первой пептидной связи до присоединения последней аминокислоты. На рибосоме имеется два участка для связывания двух молекул т-РНК. В одном участке-пептидильном(П) находится первая т-РНК с аминокислотой метионин и с него начинается синтез любой молекулы белка. Во второй участок рибосомы- аминоацильный (А) поступает вторая молекула т-РНК и присоединяется к своему кодону. Между метионином и второй аминокислотой образуется пептидная связь. Вторая т-РНК перемещается вместе со своим кодоном м-РНК в пептидильный центр. Перемещение т-РНК с полипептидной цепочкой из аминоацильного центра в пептидильный сопровождается продвижением рибосомы по м-РНК на шаг, соответствующий одному кодону. Т-РНК, доставившая метионин, возвращается в цитоплазму, амноацильный центр освобождается. В него поступает новая т-РНК с аминокислотой, зашифрованной очередным кодоном. Между третьей и второй аминокислотами образуется пептидная связь и третья т-РНК вместе с кодоном м-РНК перемещается в пептидильный центр.Процесс элонгации, удлинения белковой цепи. Продолжается до тех пор, пока в рибосому не попадёт один из трёх кодонов, не кодирующих аминокислоты. Это кодон — терминатор и для него не существует соответствущей т-РНК, поэтому ни одна из т-РНК не может занять место в аминоацильном центре.
Терминация
– завершение синтеза полипептида. Она связана с узнаванием специфическим рибосомным белком одного из терминирующих кодонов (УАА, УАГ, УГА), когда он будет входить в аминоацильный центр. К рибосоме присоединяется специальный фактор терминации, который способствует разъединению субъединиц рибосомы и освобождению синтезированной молекулы белка. К последней аминокислоте пептида присоединяется вода и её карбоксильный конец отделяется от т-РНК.
Сборка пептидной цепи осуществляется с большой скоростью. У бактерий при температуре 37°С она выражается в добавлении к полипептиду от 12 до 17 аминокислот в секунду. В эукариотических клетках к полипептиду добавляется две аминокислоты в одну секунду.
Синтезированная полипептидная цепь затем поступает в комплекс Гольджи, где завершается построение белковой молекулы (последовательно возникают вторая, третья, четвертая структуры). Здесь же происходит комплексование белковых молекул с жирами и углеводами.
Весь процесс биосинтеза белка представлен в виде схемы: ДНК ® про иРНК ® мРНК ® полипептидная цепь ® белок® комплексование белков и их преобразование в функционально активные молекулы.
Этапы реализации наследственной информации также протекают сходным образом: сначала она транскрибируется в нуклеотидную последовательность мРНК, а затем транслируется в аминокислотную последовательность полипептида на рибосомах с участием тРНК.
Транскрипция эукариот осуществляется под действием трех ядерных РНК-полимераз. РНК-полимераза 1 находится в ядрышках и отвечает за транскрипцию генов рРНК. РНК-полимераза 2 находится в ядерном соке и отвечает за синтез предшественника мРНК. РНК-полимераза 3 –небольшая фракция в ядерном соке, которая осуществляет синтез малых рРНК и тРНК. РНК-полимеразы специфически узнают нуклеотидную последовательность транскрипции-промотор. Эукариотическая мРНК вначале синтезируется в виде предшественницы (про- иРНК), на нее списывается информация с экзонов и интронов. Синтезированная мРНК обладает большими, чем необходимо для трансляции размерами и оказывается менее стабильной.
В процессе созревания молекулы мРНК с помощью ферментов рестриктаз вырезаются интроны, а с помощью ферментов – лигаз сшиваются экзоны. Созревание мРНК называется процессингом, сшивание экзонов называется сплайсингом. Таким образом, зрелая мРНК содержит только экзоны и она значительно короче её предшественницы – про- иРНК. Размеры интронов варьируют от 100 до 10000 нуклеотидов и более. На долю интонов приходится около 80% всей незрелой мРНК. В настоящее время доказана возможность альтернативного сплайсинга, при котором из одного первичного транскрипта могут удаляться в разных его участках нуклеотидные последовательности и будут образовываться несколько зрелых мРНК. Данный вид сплайсинга характерен в системе генов иммуноглобулинов у млекопитающих, что даёт возможность формировать на основе одного транскрипта мРНК разные виды антител. По завершению процессинга зрелая мРНК проходит отбор перед выходом в цитоплазму из ядра. Установлено, что попадает всего 5% зрелой мРНК, а остальная часть расщепляется в ядре. Преобразование первичных транскриптонов эукариотических генов, связанное с их экзон-интронной организацией, и в связи с переходом зрелой мРНК из ядра в цитоплазму, определяет особенности реализации генетической информации эукариот. Следовательно, мозаичный ген эукариот не является геном цистроном, так как не вся последовательность ДНК используется для синтеза белка.
Процесс биосинтеза белка осуществляется на рибосомах, а хранителем генетической информации является ДНК- Для передачи информации с ДНК, нахо -дящейся в ядре, к месту синтеза белка требуется посредник. Его роль выполняет информационная (матричная) РНК, которая синтезируется на одной из цепей молекулы ДНК по принципу комплементарности.
Таким образом, реализация наследственной информации в клетке осуществляется в два этапа: сначала информация о структуре белка копируется с ДНК на иРНК (транскрипция), а затем реализуется на рибосоме в виде конечного продукта — белка (трансляция). Это можно представить в виде схемы:
Транскрипция. Переписывание наследственной информации с ДНК на иРНК называется транскрипцией (от лат. транскрипцио — переписывание). Этот процесс происходит следующим образом.
На определенном участке молекулы ДНК происходит разъединение комплементарных цепей. Вдоль одной из цепей (ее называют транскрибируемой цепью) начинает движение фермент РНК-полимераза.
в) генетический код
РНК-полимераза синтезирует из нуклеотидов молекулу иРНК, при этом транскрибируемая цепь ДНК используется в качестве матрицы (рис. 65). Полученная иРНК комплементарна участку транскрибируемой цепи ДНК, значит, порядок нуклеотидов в иРНК строго определен порядком нуклеотидов в ДНК Например, если участок транскрибируемой цепи ДНК имеет последовательность нуклеотидов А Ц Г Т Г А, то соответствующий участок молекулы иРНК будет иметь вид У Г ЦАЦУ (обратите внимание, что в состав нуклеотидов РНК вместо тимина входит урацил). Таким образом, в результате транскрипции генетическая информация переписывается с ДНК на иРНК
Транскрипция может происходить одновременно на нескольких генах одной хромосомы и на генах, расположенных в разных хромосомах.
Поскольку в одной молекуле ДНК содержится множество генов, очень важно, чтобы РНК-полимераза начинала синтез иРНК со строго определенного участка ДНК- Поэтому в начале каждого гена находится особая специфическая последовательность нуклеотидов, называемая промотором. РНК-полимераза распознает промотор, взаимодействует с ним и начинает синтез цепочки иРНК с нужного места. Фермент синтезирует иРНК, присоединяя к ней новые нуклеотиды, пока не дойдет до особой последовательности нуклеотидов в молекуле ДНК — терминатора. Эта последовательность нуклеотидов указывает на то, что синтез иРНК нужно прекратить.
У прокариот синтезированные молекулы иРНК могут сразу же взаимодействовать с рибосомами и участвовать в синтезе белков. У эукариот иРНК синтезируется в ядре. Там она взаимодействует со специальными ядерными белками и переносится через поры в ядерной мембране в цитоплазму.
На специальных генах синтезируются и два других типа РНК: тРНК и рРНК
Трансляция. Процесс синтеза белка из аминокислот, происходящий на рибосомах, называется трансляцией (от лат. трансляцио — перевод). В ходе трансляции осуществляется перевод последовательности нуклеотидов молекулы иРНК в последовательность аминокислот молекулы белка. Иными словами, «язык» нуклеотидов переводится на «язык» аминокислот.
В цитоплазме обязательно должен быть полный набор аминокислот, необходимых для синтеза белков. Эти аминокислоты образуются в результате расщепления белков, получаемых организмом с пищей, или синтезируются в самом организме.
Информационная РНК связывается с малой субъединицей рибосомы, после этого присоединяется большая субъединица (рис. 66).
Синтез белка начинается со стартового кодона АУТ. Так как этот триплет кодирует аминокислоту метионин, то все белки (за исключением особых случаев) будут начинаться с остатка метионина. Отщепление этого остатка у большинства белков происходит позднее, в ходе созревания белковой молекулы.
Начиная со стартового кодона, молекула иРНК последовательно, триплет за триплетом, продвигается через рибосому, что сопровождается ростом полипеп-тидной цепочки. Соединение аминокислот в нужную последовательность (в соответствии с кодонами иРНК) осуществляется на рибосомах при участии транспортных р н к
Благодаря специфическому расположению комплементарных нуклеотидов молекула тРНК, как уже отмечалось, имеет форму, напоминающую лист клевера (рис. 67). У каждой тРНК имеется акцепторный конец, к которому присоединяется определенная аминокислота, предварительно активированная энергией АТФ. Для активации одной аминокислоты необходимо расщепить одну молекулу АТФ.
В противоположной части молекулы тРНК находится специфический триплет — ант и кодон, ответственный за прикрепление по принципу комплемен-тарности к соответствующему триплету иРНК (кодону).
Молекула тРНК с присоединенной активированной аминокислотой благодаря антикодону комплементарно связывается с соответствующим кодоном иРНК Таким же образом к следующему кодону иРНК прикрепляется вторая тРНК с активированной аминокислотой. Между двумя аминокислотами возникает пептидная связь, после чего первая тРНК освобождается от аминокислоты и покидает рибосому.

После этого иРНК сдвигается на один триплет, а в рибосому проникает следующая молекула тРНК с аминокислотой. В результате к образованному дипептиду присоединяется третья аминокислота и иРНК сдвигается еще на один триплет. Так происходит наращивание полипептид-ной цепочки.
Процесс трансляции продолжается до тех пор, пока в рибосому не попадет один из трех стоп-кодонов:

УАА, УАГ или УГА, после чего синтез белка прекращается и рибосома распадается на две субъединицы.
Все описанные реакции происходят очень быстро. Подсчитано, что синтез крупной молекулы белка осуществляется приблизительно за 1 —2 мин.
Каждый этап биосинтеза белка катализируется соответствующими ферментами и снабжается энергией за счет расщепления АТФ.
Молекула иРНК может связываться одновременно с несколькими рибосомами. Комплекс из иРНК и рибосом (от 5—6 до нескольких десятков) называется пол и сомой. Образование полисом повышает эффективность функционирования иРНК, так как позволяет одновременно осуществлять синтез нескольких одинаковых молекул белка.
Если синтез белка происходил на рибосомах, связанных с шероховатой ЭПС, то образовавшаяся полипептидная цепь сначала оказывается внутри полости эндоплазматической сети, а затем транспортируется в комплекс Гольджи. В этих органоидах происходит созревание белка — формирование вторичной, третичной и четвертичной структуры, присоединение к белковой молекуле небелковых компонентов и др. Если синтез белка осуществлялся на свободных рибосомах, расположенных в гиалоплазме, то синтезированная белковая молекула транспортируется в нужную часть клетки, где и приобретает соответствующую структуру.
Таким образом, генетическая информация, которая содержится в ДНК, в результате процессов транскрипции и трансляции реализуется в клетке в виде молекул белков. Синтез белка обеспечивается взаимодействием всех типов РНК: рРНК является главным структурным компонентом рибосом, иРНК — носителем информации о первичной структуре белка, тРНК доставляют на рибосому аминокислоты, а также обеспечивают их правильное включение в полипептид-ную цепь.
Биосинтез РНК (транскрипция) и биосинтез белка (трансляция) осуществляются с использованием матриц — ДНК и иРНК соответственно. Поэтому, так же как и репликация, процессы транскрипции и трансляции являются реакциями матричного синтеза.
1. Какие процессы относятся к реакциям матричного синтеза?
Брожение, трансляция, транскрипция, фотосинтез, репликация.
2. Что такое транскрипция? Как протекает этот процесс?
3. Какой процесс называется трансляцией? Охарактеризуйте основные этапы трансляции.
4. Почему при трансляции в состав белка включаются не любые аминокислоты в случайном порядке, а только те, которые закодированы триплетами иРНК, причем в строгом соответствии с последовательностью этих триплетов? Как вы думаете, сколько видов тРНК участвует в синтезе белков в клетке?
5. Реакции матричного синтеза следует относить к процессам ассимиляции или диссимиляции? Почему?
6. Участок транскрибируемой цепи ДНК имеет следующий порядок нуклеотидов: ТАЦТГГАЦАТАТТАЦААГАЦТ. Установите последовательность аминокислотных остатков пептида, закодированного этим участком.
7. Исследования показали, что в молекуле иРНК 34% от общего числа азотистых оснований приходится на гуанин, 18 % — на урацил, 28 % — на цитозин и 20 % — на аденин. Определите процентный состав азотистых оснований двуцепочечного участка ДНК, одна из цепей которого служила матрицей для синтеза данной иРНК.
8. В эритроцитах млекопитающих синтез гемоглобина может происходить еще в течение нескольких дней после утраты этими клетками ядер. Как вы можете это объяснить?
- § 1. Содержание химических элементов в организме. Макро- и микроэлементы
- § 2. Химические соединения в живых организмах. Неорганические вещества
- § 10. История открытия клетки. Создание клеточной теории
- § 15. Эндоплазматическая сеть. Комплекс Гольджи. Лизосомы
- § 24. Общая характеристика обмена веществ и преобразование энергии
Глава 1. Химические компоненты живых организмов
Глава 2. Клетка - структурная и функциональная единица живых организмов
Глава 3. Обмен веществ и преобразование энергии в организме
Глава 4. Структурная организация и регуляция функций в живых организмах
После открытия принципа молекулярной организации такого вещества, как ДНК в 1953 году, начала развиваться молекулярная биология. Далее в процессе исследований ученые выяснили как рекомбенируется ДНК, ее состав и как устроен наш человеческий геном.
Каждый день на молекулярном уровне происходят сложнейшие процессы. Как устроена молекула ДНК, из чего она состоит? И какую роль играют в клетке молекулы ДНК? Расскажем подробно обо всех процессах, происходящих внутри двойной цепи.
Что такое наследственная информация?
Итак, с чего все начиналось? Еще в 1868 нашли в ядрах бактерий. А в 1928 г. Н. Кольцов выдвинул теорию о том, что именно в ДНК зашифрована вся генетическая информация о живом организме. Затем Дж. Уотсон и Ф. Крик нашли модель всем теперь известной спирали ДНК в 1953 году, за что заслужено получили признание и награду — Нобелевскую премию.
Что такое вообще ДНК? Это вещество состоит из 2 объединенных нитей, точнее спиралей. Участок такой цепочки с определенной информацией называется геном.

В ДНК хранится вся информация о том, что за белки будут формироваться и в каком порядке. Макромолекула ДНК — это материальный носитель невероятно объемной информации, которая записана строгой последовательностью отдельных кирпичиков — нуклеотидов. Всего нуклеотидов 4, они дополняют друг друга химически и геометрически. Этот принцип дополнения, или комплементарности, в науке будет описан позже. Это правило играет ключевую роль в кодировке и декодировании генетической информации.
Так как нить ДНК невероятно длинная, повторений в этой последовательности не бывает. У каждого живого существа собственная уникальная цепочка ДНК.
Функции ДНК
К функциям относятся хранение наследственной информации и ее
передача потомству. Без этой функции геном вида не мог бы сохраняться и развиваться на протяжении тысячелетий.
Организмы, которые претерпели серьезные
мутации генов, чаще не выживают или теряют способность производить потомство. Так происходит природная защита от вырождения вида.

Еще одна существенно важная функция — реализация хранимой информации. Клетка не может создать ни одного жизненно важного белка без тех инструкций, которые хранятся в двойной цепочке.
Состав нуклеиновых кислот
Сейчас уже достоверно известно, из чего состоят сами нуклеотиды — кирпичики ДНК. В их состав входят 3 вещества:
- Ортофосфорная кислота.
- Азотистое основание. Пиримидиновые основания — которые имеют только одно кольцо. К ним относят тимин и цитозин. Пуриновые основания, в составе которых присутствуют 2 кольца. Это гуанин и аденин.
- Сахароза. В составе ДНК — дезоксирибоза, В РНК — рибоза.
Число нуклеотидов всегда равно числу азотистых оснований. В специальных лабораториях расщепляют нуклеотид и выделяют из него азотистое основание. Так изучают отдельные свойства этих нуклеотидов и возможные мутации в них.
Уровни организации наследственной информации
Разделяют 3 уровня организации: генный, хромосомный и геномный. Вся информация, нужная для синтеза нового белка, содержится на небольшом участке цепочки — гене. То есть ген считается низший и самый простой уровень кодировки информации.

Гены, в свою очередь, собраны в хромосомы. Благодаря такой организации носителя наследственного материала группы признаков по определенным законам чередуются и передаются от одного поколения к другому. Надо заметить, генов в организме невероятно много, но информация не теряется, даже когда много раз рекомбенируется.
Разделяют несколько видов генов:
- по функциональному назначению выделяют 2 типа: структурные и регуляторные последовательности;
- по влиянию на процессы, протекающие в клетке, различают: супервитальные, летальные, условно летальные гены, а также гены мутаторы и антимутаторы.
Располагаются гены вдоль хромосомы в линейном порядке. В хромосомах информация сфокусирована не вразброс, существует определенный порядок. Существует даже карта, в которой отображены позиции, или локусы генов. Например, известно, что в хромосоме № 18 зашифрованы данные о цвете глаз ребенка .
А что же такое геном? Так называют всю совокупность нуклеотидных последовательностей в клетке организма. Геном характеризует целый вид, а не отдельную особь.
Каков генетический код человека?
Дело в том, что весь огромнейший потенциал человеческого развития заложен уже в период зачатия. Вся наследственная информация, которая необходима для развития зиготы и роста ребенка уже после рождения, зашифрована в генах. Участки ДНК и есть самые основные носители наследственной информации.
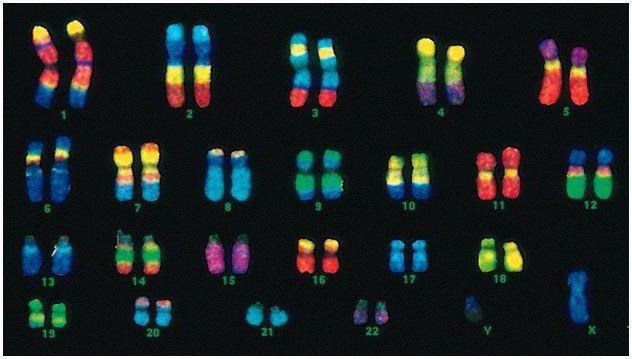
У человека 46 хромосом, или 22 соматические пары плюс по одной определяющей пол хромосоме от каждого родителя. Этот диплоидный набор хромосом кодирует весь физический облик человека, его умственные и физические способности и предрасположенность к заболеваниям. Соматические хромосомы внешне неразличимы, но несут они разную информацию, так как одна из них от отца, другая - от матери.
Мужской код отличается от женского последней парой хромосом — ХУ. Женский диплоидный набор — это последняя пара, ХХ. Мужчинам достается одна Х-хромосома от биологической матери, и затем она передается дочерям. Половая У-хромосома передается сыновьям.
Хромосомы человека значительно разнятся по размеру. Например, самая маленькая пара хромосом - №17. А самая большая пара - 1 и 3.
Диаметр двойной спирали у человека - всего 2 нм. ДНК настолько плотно закручена, что вмещается в маленьком ядре клетки, хотя ее длина будет достигать 2 метров, если ее раскрутить. Длина спирали — это сотни миллионов нуклеотидов.
Как передается генетический код?
Итак, какую роль играют в клетке молекулы ДНК при делении? Гены — носители наследственной информации - находятся внутри каждой клетки организма. Чтобы передать свой код дочернему организму, многие существа делят свое ДНК на 2 одинаковые спирали. Это называется репликацией. В процессе репликации ДНК расплетается и специальные «машины» дополняют каждую цепочку. После того как раздвоится генетическая спираль, начинает делиться ядро и все органеллы, а затем и вся клетка.
Но у человека другой процесс передачи генов - половой. Признаки отца и матери перемешиваются, в новом генетическом коде содержится информация от обоих родителей.
Хранение и передача наследственной информации возможны благодаря сложной организации спирали ДНК. Ведь как мы говорили, структура белков зашифрована именно в генах. Раз создавшись во время зачатия, этот код на протяжении всей жизни будет копировать сам себя. Кариотип (личный набор хромосом) не изменяется во время обновления клеток органов. Передача же информации осуществляется с помощью половых гамет — мужских и женских.
Передавать свою информацию потомству не способны только вирусы, содержащие одну цепочку РНК. Поэтому, чтобы воспроизводиться, им нужны клетки человека или животного.
Реализация наследственной информации
В ядре клетки постоянно происходят важные процессы. Вся информация, записанная в хромосомах, используется для построения белков из аминокислот. Но цепочка ДНК никогда не покидает ядро, поэтому здесь нужна помощь другого важного соединения = РНК. Как раз РНК способно проникнуть через мембрану ядра и взаимодействовать с цепочкой ДНК.
Посредством взаимодействия ДНК и 3 видов РНК происходит реализация всей закодированной информации. На каком уровне происходит реализация наследственной информации? Все взаимодействия происходят на уровне нуклеотидов. Информационная РНК копирует участок цепи ДНК и приносит эту копию в рибосому. Здесь начинается синтез из нуклеотидов новой молекулы.
Для того чтобы иРНК могла скопировать необходимую часть цепи, спираль разворачивается, а затем, по завершении процесса перекодировки, снова восстанавливается. Причем этот процесс может происходить одновременно на 2 сторонах 1 хромосомы.
Принцип комплементарности
Состоят из 4 нуклеотидов — это аденин (А), гуанин (G), цитозин (С), тимин (T). Соединены они водородными связями по правилу комплементарности. Работы Э. Чаргаффа помогли установить это правило, так как ученый заметил некоторые закономерности в поведении этих веществ. Э. Чаргафф открыл, что молярное отношение аденина к тимину равно единице. И точно так же отношение гуанина к цитозину всегда равно единице.
На основе его работ генетики сформировали правило взаимодействия нуклеотидов. Правило комплементарности гласит, что аденин соединяется только с тимином, а гуанин - с цитозином. Во время декодирования спирали и синтеза нового белка в рибосоме такое правило чередования помогает быстро найти необходимую аминокислоту, которая прикреплена к транспортной РНК.
РНК и его виды
Что такое наследственная информация? нуклеотидов в двойной цепи ДНК. А что такое РНК? В чем заключается ее работа? РНК, или рибонуклеиновая кислота, помогает извлекать информацию из ДНК, декодировать ее и на основе принципа комплементарности создавать необходимые клеткам белки.
Всего выделяют 3 вида РНК. Каждая из них выполняет строго свою функцию.
- Информационная (иРНК) , или еще ее называют матричная. Она заходит прямо в центр клетки, в ядро. Находит в одной из хромосом необходимый генетический материал для постройки белка и копирует одну из сторон двойной цепи. Копирование происходит снова по принципу комплементарности.
- Транспортная — это небольшая молекула, у которой на одной стороне декодеры-нуклеотиды, а на другой стороне соответствующие основному коду аминокислоты. Задача тРНК — доставить в «цех», то есть в рибосому, где синтезирует необходимую аминокислоту.
- рРНК — рибосомная. Она контролирует количество белка, который продуцируется. Состоит из 2 частей — аминокислотного и пептидного участка.
Единственное отличие при декодировании — у РНК нет тимина. Вместо тимина тут присутствует урацил. Но потом, в процессе синтеза белка, при ТРНК все равно правильно устанавливает все аминокислоты. Если же происходят какие-то сбои в декодировании информации, то возникает мутация.
Репарация поврежденной молекулы ДНК
Процесс восстановления поврежденной двойной цепочки называется репарацией. В процессе репарации поврежденные гены удаляются.

Затем необходимая последовательность элементов в точности воспроизводиться и врезается обратно в то же место на цепи, откуда было извлечено. Все это происходит благодаря специальным химическим веществам — ферментам.
Почему происходят мутации?
Почему некоторые гены начинают мутировать и перестают выполнять свою функцию — хранение жизненно необходимой наследственной информации? Это происходит из-за ошибки при декодировании. Например, если аденин случайно заменен на тимин.
Существуют также хромосомные и геномные мутации. Хромосомные мутации случаются, если участки наследственной информации выпадают, удваиваются либо вообще переносятся и встраиваются в другую хромосому.

Геномные мутации наиболее серьезны . Их причина - это изменение числа хромосом. То есть когда вместо пары — диплоидного набора присутствует в кариотипе триплоидный набор.
Наиболее известный пример триплоидной мутации — это синдром Дауна, при котором личный набор хромосом 47. У таких детей образуется 3 хромосомы на месте 21-й пары.
Известна также такая мутация, как полиплодия. Но полиплодия встречается только у растений.


