Клітинна стінка з целюлози в когось. Клітинна стінка
Клітинна стінка(Клітинна оболонка)- Характерна ознака рослинної клітини, що відрізняє її від клітини тварини. Клітинна стінка надає клітині певної форми. Культивовані на спеціальних живильних середовищах клітини рослин, у яких ферментативним шляхом видаляється стінка, завжди набувають сферичної форми. Клітинна стінка надає клітині міцності та захищає протопласт, вона врівноважує тургорний тиск і перешкоджає, таким чином, розриву плазмалеми. Сукупність клітинних стін утворює внутрішній скелет, що підтримує тіло рослини і надає йому механічної міцності.
Майже у всіх пологів бактерій є одна, яка є жорсткою структурою, яка містить вуглеводи, які оточують бактеріальну клітину. Цей екзоскелет клітинної стінки дає бактеріям кілька переваг. Клітинна стінка захищає бактерії від ушкоджень, оточуючи її жорсткою та жорсткою структурою. Ця структура також пориста. Малі молекули можуть вільно проходити через клітинну стінку до мембрани, але великі молекуливиключено. Таким чином, клітинна стінка діє як грубий фільтр.
Однак основна функція клітинної стінки полягає у підтримці форми комірки та запобіганні розриву осмотичного тиску. Не дивно, що природа вирішила проблему. Клітинна стінка зберігає свою форму та проникність! Копаючись у деталях, ми можемо зрозуміти, як це робить стінка клітини.
Клітинна стінка безбарвна та прозора, легко пропускає сонячне світло. Зазвичай стіни просочені водою. За системою клітинних стінок здійснюється транспорт води та розчинених у ній низькомолекулярних сполук (транспорт по апопласту).
Клітинна стінка складається в основному з полісахаридів, які можна поділити на скелетні речовиниі речовини матриксу.
Якими результатами є плоский та схрещений малюнок, який є дуже сильним та жорстким, але досить відкритим для руху частинок. Як ви можете бачити, пептидоглікан нагадує забір ланцюга. Майже всі бактерії мають клітинні стінки з пептидоглікану. Але до стінки клітини більше, ніж до пептидоглікану. У природі існують два основних типи клітинних стінок: грампозитивні та грамнегативні, кожен з дуже різними структурами. Термін "Грам" відноситься до техніки забарвлення за Грамом, яка розрізняє бактерії з двома різними стінками клітин.
Скелетною речовиноюклітинної стінки рослин є целюлоза (клітковина), Що являє собою бета-1,4-D-глюкан Це найпоширеніша органічна речовина біосфери. Молекули целюлози є дуже довгими нерозгалуженими ланцюгами, вони розташовуються паралельно один одному групами по кілька десятків і скріплені численними водневими зв'язками. В результаті утворюються мікрофібрили, які створюють структурний каркас стінки та зумовлюють її міцність. Мікрофібрили целюлози видно лише в електронному мікроскопі, їх діаметр дорівнює 10-30 нм, довжина досягає декількох мкм.
Для цього уроку не важливо знати точну процедуру фарбування. Грампозитивні бактерії мають кілька шарів пептидоглікану, що утворюють дуже товсті та жорсткі клітинні стінки. Плоскі та поперечні шари пептидоглікану укладаються один поверх іншого, створюючи відносно товсту клітинну стінку. Досягнення пептидогліканської палі - це чайна кислота. Ця довга молекула має негативний зарядта допомагає переміщати іони через товсту клітинну стінку. Важливо, що чайна кислота міститься лише в грампозитивних бактеріях.
Целюлоза нерозчинна та не набухає у воді. Вона дуже інертна у хімічному відношенні, не розчиняється в органічних розчинниках, концентрованих лугах та розведених кислотах. Мікрофібрили целюлози еластичні та дуже міцні на розрив (подібні зі сталлю). Ці властивості визначають широке застосування целюлози та її продуктів. Світова продукція бавовняного волокна, що складається майже цілком із целюлози, становить 1,5 10 7 тонн на рік. З целюлози отримують бездимний порох, ацетатний шовк та віскозу, целофан, папір. Якісну реакціюна целюлозу проводять з реактивом хлор-цинк-йод, целюлозна клітинна стінка забарвлюється у синьо-фіолетовий колір.
Грамнегативні бактерії відрізняються від грампозитивних двома основними способами. Грамнегативна клітинна стінка складається тільки з одного або двох шарів пептидоглікану, який покритий. зовнішньою мембраною. Грамнегативні бактерії мають типову клітинну мембрану, яка покриває всю клітину. Прямо за межами цієї мембрани знаходиться периплазма, яка являє собою драглистий шар між зовнішньою мембраною і клітинною мембраною. У периплазматичний простір входять пептидогліканові шари, а також ферменти та додаткові транспортні білки.
У грибів скелетною речовиною клітинної стінки є хітін- Полісахарид, побудований з залишків глюкозаміну. Хітін ще міцніший, ніж целюлоза.
Мікрофібрили занурені в аморфний матриксзазвичай являє собою насичений водою пластичний гель. Матрикс є складною сумішшю полісахаридів, молекули яких складаються з залишків декількох різних цукрів і є більш короткими, ніж у целюлози, і розгалужені ланцюги. Матричні полісахариди визначають такі властивості клітинної стінки, як сильна набухання, висока проникність для води та розчинених у ній низькомолекулярних сполук, катіонообмінні властивості. Полісахариди матриксу ділять на дві групи. пектинові речовиниі геміцелюлози.
У грамнегативних бактеріях є лише один або два шари пептидогліканової сітки. Існують чотири основні структури, що поділяються всіма прокаріотичними, бактеріальними або архаїчними клітинами. Плазмова мембрана Цитоплазма рибосоми Генетичний матеріал. . Деякі прокаріотичні клітини також мають інші структури, такі як клітинна стінка, пилки та джгутики. Кожна з цих клітинних структурі компонентів відіграє вирішальну роль у зростанні, виживанні та розмноженні прокаріотичних клітин.
Клітинна стінка грибів є динамічну структуру, яка захищає клітину від змін осмотичного тиску та інших напруг навколишнього середовища, дозволяючи клітині взаємодіяти з довкіллям. Структура і біосинтез клітинної стінки грибів є унікальними для грибів і, отже, є відмінною мішенню для розробки протигрибкових препаратів. Розглянуто структуру стінки грибкової клітини та ліків, націлених на її біосинтез.
Пектинові речовинисильно набухають чи розчиняються у воді. Вони легко руйнуються під дією лугів та кислот. Найпростішими представниками пектинових речовин є розчинні у воді пектові кислоти– продукти полімеризації альфа-D-галактуронової кислоти (до 100 одиниць), пов'язаних 1,4-зв'язками в лінійних ланцюгах (альфа-1,4-D-галактуронан). Пектинові кислоти (пектин)– це високомолекулярні (100-200 одиниць) полімерні сполуки альфа-D-галактуронової кислоти, в яких карбоксильні групи частково метильовані. Пектатиі пектинати– кальцієві та магнієві солі пектових та пектинових кислот. Пектинові кислоти, пектати та пектинати розчиняються у воді в присутності цукрів та органічних кислот з утворенням щільних гелів.
На основі досліджень кількох грибів було показано, що клітинна стінка складається в основному з хітину, глюканів, маннанів та глікопротеїнів. Обговорюється біосинтез різних компонентівклітинної стінки грибів та важливість компонентів у формуванні функціональної клітинної стінки, що показано в результаті мутаційних аналізів. Є переконливі докази того, що хітин, глюкани та глікопротеїни ковалентно пошиті і що зшивання є динамічний процесщо відбувається позаклітинно.
Ми витратили багато часу на те, що знаходиться всередині комірки, але що знаходиться за її межами? Це залежить від того, який тип осередку задіяний. Клітини рослин і грибів мають жорстку клітинну стінку, яка захищає та підтримує їх, тоді як клітини тварин виділяють матеріали навколо них, утворюючи мережу макромолекул, відому як позаклітинний матрикс. Тут ми побачимо більш детально ці зовнішні структури та функції, які вони виконують у різних типахосередків.
У клітинних стінках рослин переважно присутні протопектини– високомолекулярні полімери метоксильованої полігалактуронової кислоти з арабінанами та галактанами, у дводольних рослин до складу ланцюгів галактуронану входить не велика кількістьрамнози. Протопектини нерозчинні у воді.
Геміцелюлозиє розгалужені ланцюги, побудовані з залишків нейтральних цукрів, частіше зустрічаються глюкоза, галактоза, манноза, ксилоза; ступінь полімеризації 50-300. Геміцелюлози хімічно стійкіші, ніж пектинові речовини, вони важче гідролізуються і слабше набухають у воді. Геміцелюлози можуть відкладатися в стінках клітин насіння як запасні речовини (фінікова пальма, хурма). Пектинові речовини та геміцелюлози пов'язані взаємними переходами. Крім полісахаридів, в матриксі клітинних стін присутній спеціальний структурний білок. Він пов'язаний із залишками цукру арабінози і тому є глікопротеїдом.
Позаклітинна матриця тварин клітин
Більшість клітин тварин виділяють матеріали у позаклітинний простір, створюючи складну мережу білків та вуглеводів, яка називається позаклітинним матриксом. Основним компонентом позаклітинного матриксу є колагеновий білок. Колагени модифікуються вуглеводами і, після секреції поза клітиною, утворюють довгі волокна, звані 1 колагеновими фібрилами.
Колаген відіграє фундаментальну роль у опорі та структурній цілісності тканин. Генетичні порушеннялюдини, які впливають на колаген, такі як синдром Елерса-Данлоса, виробляють тендітні тканини, які надто легко розтягуються та розриваються. 2 2 2 починають верхній індекс, 2, кінець верхнього індексу.
Матричні полісахариди не просто заповнюють проміжки між целюлозними мікрофібрилами. Їхні ланцюги розташовуються впорядковано і утворюють численні зв'язки як один з одним, так і з мікрофібрил, що значно підвищує міцність клітинної стінки.
Клітинні стінки рослин часто піддаються хімічним змінам. Одервеснення, або лігніфікація відбувається у тому випадку, якщо в матриксі відкладається лігнін– полімерна сполука фенольної природи, нерозчинна у воді. Клітинна стінка, що здеревніла, втрачає еластичність, різко підвищується її твердість і міцність на стиск, знижується проникність для води. Реактивами на лігнін є: 1) флороглюцині концентрована хлористоводневаабо сірчана кислота(одревеснілі стінки набувають вишнево-червоного забарвлення) і 2) сульфат аніліну, під впливом якого здерев'янілі стіни стають лимонно-жовтими. Лігніфікація характерна для стінок клітин провідної тканини ксилеми (деревини) та механічної тканини склеренхіми.
У позаклітинній матриці волокна колагену переплітаються з класом протеогліканів, які мають вуглеводи, прикріплені до них, такі як скелет полісахариду, показаний на зображенні нижче. Позаклітинна матриця містить інші типи білків і вуглеводів.
Протеоглікани є особливий клас глікопротеїнів з певними відмінними характеристиками. Вони дуже глікозильовані, тобто мають високий вміст вуглеводів, залежно від їх вмісту білка. Крім того, вуглеводні ланцюги протеогліканів містять специфічні цукру з сульфатними модифікаціями, пов'язаними разом з утворенням лінійних ланцюгів 1 1 1, верхній верхній індекс, 1, кінцевий верхній індекс.
Опробковування, або суберинізаціявідбувається в результаті відкладення з внутрішньої сторони клітинної стінки гідрофобних полімерів – Суберінаі воску. Суберин являє собою суміш ефірів полімерних жирних кислот. Мономерами воску є жирні спирти та воскові ефіри. Віск легко витягується органічними розчинниками та швидко плавиться, утворює кристали. Суберин – аморфна сполука, яка не плавиться і не розчиняється в органічних розчинниках. Суберин і віск, утворюючи паралельні шари, що чергуються, вистилають всю порожнину клітини з внутрішньої сторони у вигляді плівки. Суберинова плівка практично непроникна для води та для газів, тому після її утворення клітина зазвичай відмирає. Опробковування притаманно стінок клітин покривної тканини пробки. Реактивом на випробувану клітинну стінку є суданIII, забарвлення оранжево-червоне.
Щоб дізнатися більше про біохімію та структуру протеогліканів, ви можете звернутися до джерела, згаданого вище. Діаграма, що показує позаклітинний матрикс та його з'єднання з коміркою. Мережа клітин колагену та протеогліканів знаходиться поза клітиною. Колаген зв'язується з білками інтегрину в плазматичній мембрані за допомогою фібронектину. Усередині клітини інтегрини з'єднуються з мікрофіламентами цитоскелету.
Позаклітинна матриця безпосередньо пов'язана з клітинами, які вона оточує. Деякі з ключових з'єднувачів - це білки, які називаються інтегринами, які вбудовані в плазматичну мембрану. Білки у позаклітинному матриксі, такі як молекули фібронектину, показані зеленим на діаграмі вище, можуть діяти як містки між інтегринами та іншими білками у позаклітинному матриксі, такому як колаген. У внутрішній частині мембрани інтегрини пов'язані з цитоскелетом.
Кутинізаціїпіддаються зовнішні стінки клітин покривної тканини епідерми. Кутіні вісквідкладаються шарами, що чергуються на зовнішній поверхні клітинної стінки у вигляді плівки – кутикули. Кутин являє собою жироподібне полімерне з'єднання, близьке по хімічної природита властивостям суберину. Кутикула оберігає рослину від зайвого випаровування води із поверхні рослини. Забарвити її можна реактивом суданIIIв оранжево-червоний колір.
Вони також дозволяють вам сприймати ваше оточення. Коагуляція крові є ще одним прикладом зв'язку між клітинами та позаклітинною матрицею. Коли клітини, що вистилають кровоносну судину, ушкоджуються, вони вивільняють рецепторний білок, який називається тканинним фактором. Коли тканинний фактор зв'язується з молекулою, яка є у позаклітинному матриксі, він викликає серію відповідей, які зменшують втрату крові. Наприклад, це змушує тромбоцити прикріплювати до стіни пошкоджену кровоносну судину та стимулює їх до утворення факторів згортання крові.
Мінералізація клітинної стінки відбувається внаслідок відкладення в матриксі великої кількості мінеральних речовин, найчастіше кремнезему (оксиду кремнію), рідше оксалату та карбонату кальцію. Мінеральні речовини надають стінці твердості та крихкості. Відкладення кремнезему характерне для клітин епідерми хвощів, осок та злаків. Придбана в результаті відокремлення жорсткість стебел і листя служить захисним засобом проти равликів, а також значно знижує поїдання та кормову цінність рослин.
Хоча рослини не виробляють колаген, вони мають свою позаклітинну опорну структуру: клітинну стінку. Клітинна стінка є жорсткою кришкою, яка оточує комірку, захищає її і надає форму і підтримку. Ви помітили, що сирі овочі, такі як селера, тріскаються, коли ви кусаєте? Це значною мірою пов'язане із жорсткістю його клітинних стінок.
Як і позаклітинний матрикс тварин, клітинна стінка рослини складається з молекул, що секретуються клітиною. Основною органічною молекулою клітинної стінки рослини є целюлоза, полісахарид, що складається з одиниць глюкози. Целюлоза утворює волокна, які називаються мікрофібрилами, такі як показані на діаграмі нижче.
У деяких спеціалізованих клітин спостерігається ослизненняклітинної стінки. При цьому замість вторинної целюлозної стінки відбувається відкладення аморфних, сильно гідратованих кислих полісахаридів у вигляді слизіві камедей, близьких за хімічною природою до пектинових речовин Слиз добре розчиняються у воді з утворенням слизових розчинів. Камеді клейкі, витягуються у нитки. У сухому вигляді вони мають рогову консистенцію. При відкладенні слизу протопласт поступово відтісняється до центру клітини, його обсяг та обсяг вакуолі поступово зменшуються. Зрештою, порожнина клітини може повністю заповнитися слизом і клітина відмирає. У деяких випадках слиз може проходити через первинну стінку клітини на поверхню. У синтезі та секреції слизу основну участь бере апарат Гольджі.
Більшість клітинних стінок містять кілька різних полісахаридів та білків. На додаток до целюлози, інші полісахариди, які зазвичай зустрічаються в клітинній стінці, являють собою геміцелюлозу та пектин, які показані на діаграмі вище. Клітинна стінка є елементом зовнішньої клітини, який ми знаходимо в різних групах з різним еволюційним походженням. Це можна спостерігати як у грибах, так і в овочах, як у бактеріях. Склад стінки рослини помітно змінюється в залежності від груп, ви можете дізнатися більше про склад клітинної стінки овочів.
Слиз, що виділяється рослинними клітинами, виконує різні функції. Так, слиз кореневого чохлика служить як мастило, що полегшує зростання кінчика кореня в грунті. Слизові залозки комахоїдних рослин (росянка) виділяють ловчий слиз, до якого приклеюються комахи. Слиз, що виділяється зовнішніми клітинами насіннєвої шкірки (льон, айва, подорожники), закріплює насіння на поверхні ґрунту та захищає проросток від висихання. Слиз забарвлюється реактивом метиленовий синійу синій колір.
Клітинна стінка в овочах поділена на 3 шари, кожна з яких має різні функції. Ці шари є: з комірки поза: вторинна стінка, яка може бути або не бути, первинна стіна, яка завжди присутня, і середня ламела, яка згодом стає більш дифузною, оскільки первинна стінка клітин є сусідніми контактами. Ви можете прочитати про внутрішню структуру цих верств у статті, яку ми присвячуємо.
Найбільш очевидною функцією стінки комірки є контроль форми комірки. Рослини, такі як сидячі та світлочутливі організми, повинні зростати у напрямку світла, тому вони повинні часто виступати проти сили тяжіння, оскільки для цього їм потрібна жорстка структураяка дозволяє їм залишатися вертикально, не споживаючи енергію. У цьому аспекті клітинна стінка діє як екзоскелет, який утримує стебла прямостоячими. Решітка целюлозних волокон біля основи дерева має бути достатньо сильною, щоб підтримувати тонни, які може важити тисячолітнє дерево.
Виділення камедей зазвичай відбувається при пораненні рослин. Наприклад, камедетечення з поранених ділянок стовбурів та гілок часто спостерігається у вишні та сливи. Вишневий клей є застиглою камедь. Камедь виконує захисну функцію, закриваючи рану із поверхні. Утворюються камеді в основному у деревних рослин із сімейств бобових (акації, трагакантові астрагали) та розоцвітих підродини сливових (вишня, зливу, абрикос). Камеді та слизу використовуються в медицині.
Але це не лише регулює зростання рослин. Клітинна стінка є морфогенним контролером рослини. Через жорсткість клітинної стінки запобігає бічне переміщення клітин або їх відокремлення один від одного. Обмеження руху, що надає стінам весь розвиток та спосіб життя рослин. Але це не просто підтримка. Клітинна стінка дозволяє розвинути великий тиск у рослинних клітинах, що є фундаментальним для водних відносин із навколишнім середовищем. Діючи як капіляри, у яких вода випаровується у верхній частині, забезпечується доступ води через нижню частину.
Клітинна стінка є продуктом життєдіяльності протопласту. Полісахариди матриксу, глікопротеїд стінки, лігнін та слизу утворюються в апараті Гольджі. Синтез целюлози, освіта та орієнтація мікрофібрил здійснюються плазмалемою. Велика роль в орієнтації мікрофібрил належить мікротрубочкам, які розташовуються мікрофібрил, що паралельно відкладаються, поблизу плазмалеми. Якщо мікротрубочки зруйнувати, утворюються лише ізодіаметричні клітини.
Утворення клітинної стінки починається під час поділу клітини. У площині поділу утворюється клітинна платівка, єдиний шар, загальний двох дочірніх клітин. Вона складається з пектинових речовин, що мають напіврідку консистенцію; целюлоза відсутня. У дорослій клітині клітинна платівка зберігається, але зазнає змін, тому її називають серединний, або міжклітинною платівкою (міжклітинною речовиною) (Рис. 2.16). Середня платівка зазвичай дуже тонка і майже невиразна.
Відразу після утворення клітинної платівки протопласти дочірніх клітин починають відкладати власну клітинну стінку. Вона відкладається зсередини як на поверхні клітинної пластинки, так і на поверхні інших клітинних стінок, що раніше належали материнській клітині. Після поділу клітина вступає у фазу росту розтягуванням, який зумовлений інтенсивним осмотичним поглинанням клітиною води, пов'язаним із утворенням та зростанням центральної вакуолі. Тургорний тиск починає розтягувати стіну, але вона не рветься завдяки тому, що в неї постійно відкладаються нові порції мікрофібрил та речовин матриксу. Відкладення нових порцій матеріалу відбувається рівномірно по всій поверхні протопласту, тому товщина стінки клітин не зменшується.
Стінки клітин, що діляться і ростуть, називають первинними. Вони містять багато (60-90%) води. У сухій речовині переважають матричні полісахариди (60-70%), вміст целюлози не перевищує 30%, відсутня лігнін. Товщина первинної стінки дуже невелика (01-05 мкм).
Для багатьох клітин відкладення клітинної стінки припиняється одночасно із припиненням росту клітини. Такі клітини оточені тонкою первинною стінкою до кінця життя ( Рис. 2.16).
Рис. 2.16. Паренхімна клітка з первинною стінкою.
В інших клітин відкладення стінки продовжується і після досягнення клітиною остаточного розміру. При цьому товщина стінки збільшується, а об'єм, який займає порожнина клітини, скорочується. Такий процес має назву вторинного потовщеннястінки, а саму стінку називають вторинної(Рис. 2.17).
Вторинна стінка може розглядатися як додаткова, що виконує головним чином механічну опорну функцію. Саме вторинна стінка є відповідальною за властивості деревини, текстильного волокна, паперу. Вторинна стінка містить значно менше води, ніж первинна; в ній переважають мікрофібрили целюлози (40-50% від маси сухої речовини), які розташовуються паралельно один до одного. З полісахаридів матриксу характерні геміцелюлози (20-30%), пектинових речовин дуже мало. Вторинні клітинні стінки, як правило, зазнають здерев'янення. У неодревесневших вторинних стінках (луб'яні волокна льону, волоски бавовнику) вміст целюлози може досягати 95%. Великий зміст та строго впорядкована орієнтація мікрофібрил визначають високі механічні властивостівторинних стін. Часто клітини, що мають вторинну одревесневу клітинну стінку, після завершення вторинного потовщення відмирають.
Середня платівка склеює сусідні клітини. Якщо її розчинити, стінки клітин втрачають зв'язок один з одним і роз'єднуються. Цей процес називається мацерація. Досить звичайна природна мацерація, при якій пектинові речовини серединної платівки переводяться в розчинний стан за допомогою ферменту пектинази і потім вимиваються водою (перестиглі плоди груші, дині, персика, банана). Часто спостерігається часткова мацерація, при якій серединна пластинка розчиняється не по всій поверхні, а лише у кутах клітин. Внаслідок тургорного тиску сусідні клітини у цих місцях округляються, внаслідок чого утворюються міжклітинні(Рис. 2.16). Міжклітини утворюють єдину розгалужену мережу, яка заповнюється парами води та газами. Таким чином, міжклітини покращують газообмін клітин.
Характерна риса вторинної стінки – її нерівномірне відкладення поверх первинної стінки, внаслідок чого у вторинній стінці залишаються незтовщені ділянки – пори. Якщо вторинна стінка не досягає великої товщини, пори виглядають як дрібні заглиблення. У клітин із потужною вторинною стінкою пори у розрізі мають вигляд радіальних каналів, що йдуть від порожнини клітини до первинної стінки. За формою порового каналу розрізняють пори двох типів – простіі про каймлені(Рис. 2.17).
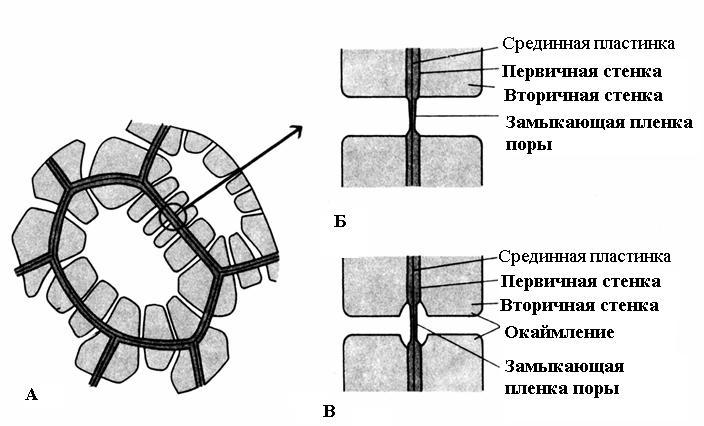
Рис. 2.17. Типи пір: А – клітини з вторинними стінками та численними простими порами; Б - пара простих пір; В – пара облямованих пір.
У простого часудіаметр порового каналу на всій довжині однаковий і має форму вузького циліндра. Прості пори характерні для паренхімних клітин, луб'яних та деревних волокон.
Пори у двох суміжних клітинах, як правило, виникають одна проти одної. Ці загальні пори мають вигляд одного каналу, розділеного тонкою перегородкою із серединної платівки та первинної стінки. Така сукупність двох пор суміжних стін сусідніх клітин носить назву пари пірта функціонує як одне ціле. Розділяючий їхній канал ділянка стінки називається замикаючою плівкою пори, або поровий мембраною. У живих клітинах замикаюча плівка пори пронизана численними плазмодесмами(Рис. 2.18).
Плазмодесмипритаманні лише рослинним клітинам. Вони є тяжі цитоплазми, що перетинають стінку суміжних клітин. Число плазмодесм в одній клітці дуже велике – від кількох сотень до десятків тисяч, зазвичай плазмодесми зібрані до груп. Діаметр плазмодесменного каналу становить 30-60 нм. Його стінки вистелені плазмалемою, безперервною з плазмалемою суміжних клітин. У центрі плазмодесми проходить мембранний циліндр. центральний стрижень плазмодесми, безперервний з мембранами елементів ендоплазматичної мережі обох клітин Між центральним стрижнем та плазмалемою в каналі знаходиться гіалоплазма, безперервна з гіалоплазмою суміжних клітин.
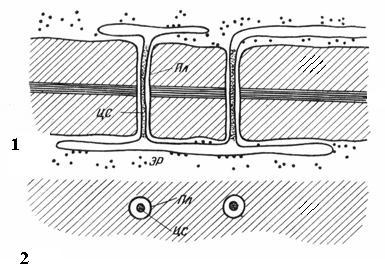
Рис. 2.18. Плазмодесми під електронним мікроскопом (схема): 1 - на поздовжньому зрізі; 2 – на поперечному зрізі; Пл- Плазмалема; ЦС – центральний стриженьплазмодесми; ЕР- Елемент ендоплазматичного ретикулуму.
Таким чином, протопласти клітин не повністю ізольовані один від одного, а повідомляються каналами плазмодесм. Ними відбувається міжклітинний транспорт іонів і дрібних молекул, і навіть передаються гормональні стимули. За допомогою плазмодесм протопласти клітин у рослинному організмі утворюють єдине ціле, зване симпластом, а транспорт речовин через плазмодесми отримав назву симпластичногона відміну від апопластичноготранспорту по клітинних стінках та міжклітинниках.
У облямованих пір(Рис. 2.17)канал різко звужується в процесі відкладення клітинної стінки, тому внутрішній отвір пори, що виходить у порожнину клітини, набагато вже, ніж зовнішнє, що упирається в первинну стінку. Облямовані пори характерні для клітин водопровідних елементів деревини, що рано відмирають. У них поровий канал у напрямку замикаючої плівки воронковидно розширюється, а вторинна стінка нависає у вигляді валика над розширеною частиною каналу, утворюючи камеру пори. Назва облямованої пори походить від того, що при розгляді з поверхні внутрішній отвір має вигляд маленького кола або вузької щілини, тоді як зовнішній отвір як би облямовує внутрішній у вигляді кола більшого діаметра або ширшої щілини.
Пори полегшують транспорт води та розчинених речовин від клітини до клітини, водночас не знижуючи міцності клітинної стінки.
механічну міцність, захищає їх вміст від пошкоджень та надмірної втрати води, підтримує форму клітин та їх розмір, а також перешкоджає розриву клітин у гіпотонічному середовищі. Клітинна стінка бере участь у поглинанні та обміні різних іонів, тобто єіонообмінником.Через клітинну оболонкуздійснюється транспорт речовин.Клітинна стінка, що формується під час поділу клітин та їх зростання шляхом розтягування, називається первинної.Після припинення зростання клітини на первинну клітинну стінку зсередини відкладаються нові шари, і утворюється міцна вториннаклітинна оболонка.
До складу клітинної стінки входять структурні компоненти(целюлоза у рослин та хітин у грибів), компоненти матриксу (геміцелюлоза, пектин, білки), інкрустуючі компоненти (лігнін, суберин) та речовини, що відкладаються на поверхні оболонки (кутин та віск).
Пектинові речовини- це полімери, побудовані з моносахаридів (арабінози та галактози), галактуронової кислоти (цукрової кислоти) та метилового спирту. Довгі молекули пектинових речовин можуть бути лінійними або розгалуженими. Молекули пектинових речовин містять велику кількість карбоксильних груп і тому здатні з'єднуватися з іонами Mg2+ та Са2-. При цьому утворюються клейкі, студнеподібні пектати магнію і кальцію, з яких потім складаються серединні пластинки, що скріплюють стінки двох сусідніх клітин.
Іони двовалентних металів можуть обмінюватися інші катіони (Н - , До + тощо. буд.). Це зумовлює катіонообмінну здатність клітинних оболонок.
Пектиновими речовинами та пектатами багаті оболонки клітин багатьох плодів. Так як при їх вилучення з оболонок і додаванні цукру утворюються гелі, пектини використовують як желе-утворювальні речовини для виготовлення мармеладу та ін.
Лігнін скріплює целюлозні волокна і діє як дуже твердий і жорсткий каркас, що підсилює міцність клітинних стінок на розтяг і стиск. Він же забезпечує клітинам додатковий захист від фізичних та хімічних впливівзнижує водопроникність. Вміст лігніну в оболонці сягає 30%. Інкрустація їм клітинних оболонок призводить до їх озеленення,яке часто спричиняє відмирання живого вмісту клітини.
Лігнін у поєднанні з целюлозою надає особливі властивостідеревини, які роблять її незамінним будівельним матеріалом.
На клітинну оболонку можуть відкладатися також жироподібні речовини - суберин, кутин і віск.
Суберінвідкладається на оболонку зсередини і робить її практично непроникною для води та розчинів. В результаті протопласт клітини відмирає та клітина заповнюється повітрям. Такий процес називається випробуванням.Спостерігається пробковування оболонки клітин у покривних тканинах багаторічних деревних рослин – перидермі, кірці, а також в ендодермі кореня.
Поверхня епідермальних клітин рослин захищена гідрофобними речовинами. кутиномі восками.Попередники цих сполук секретуються з цитоплазми на поверхню, де відбувається їх полімеризація. Шар кутину зазвичай пронизаний полісахаридними компонентами (целюлозою та пектином) і утворює кутикулу.Віск часто відкладається в кристалічній форміна поверхні частин рослин (листя, плодів), утворюючи восковий наліт.
Кутикула та восковий наліт захищають клітини від пошкоджень та проникнення інфекції, зменшують випаровування води з поверхні органів.
В оболонках епідермальних клітин деяких рослин (злаків, осок та ін.) накопичується велика кількість мінеральних речовин (Мінералізація),в першу чергу карбонату кальцію та кремнезему. При мінералізації листя і стебла рослин стають жорсткими, твердими і меншою мірою поїдаються тваринами.
Таким чином, клітинна стінка грає важливу рольу житті клітин рослин та грибів і виконує ряд специфічних функцій.
Джерело : Н.А. Лемеза Л.В.Камлюк Н.Д. Лісов "Посібник з біології для вступників до ВНЗ"


