Потовщення на стеблі, як називається. Класифікація по поперечному перерізу стебла
Округлі (наприклад: соняшник)
Чотиригранні
Тригранні
Багатогранні (ребристі) (наприклад: гарбуз)
Крилаті (мають шкірясті вирости)
Формування стебла
Стебло, як і корінь, формується із первинної верхівкової меристеми. На відміну від кореня, конус наростання захищений від довкіллямолодими листочками, що розвиваються. Як і в корені клітини меристеми діляться мітозом і спеціалізуються в первинні постійні тканини, але на відміну від кореня в стеблі немає чітко виражених зон росту. Спеціалізація клітин відбувається дуже швидко, і так само як і в корені ці клітини спеціалізуються на трьох таких же ділянках.
З периферії 1 шар клітин перетворюється на первинну покривну тканину; у первинну кору; у центрі виникає центральний осьовий циліндр. У стеблі, на відміну кореня центральний циліндр багато разів займає більше місцяніж первинна кора. 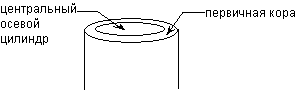
Первинна покривна тканинастебла епідерміс, 1 шар, з продихами, з волосками. Його функція – захист нижчих структур.
Первинна корастебло відповідає за фотосинтез і надає механічної стійкості (опирається від вигину). Визначає стебло у просторі. Вона складається з:
1) хлоренхіму (від 1 до безлічі шарів цих клітин)
2) коленхіму (від 1 і більше шарів). У однодольних трапляється рідко.
3) ендодерма
Ендодерма стебла є 1 шар живих клітин, з целюлозними оболонками, що містять крохмальні зерна. Вони не є запасною речовиною, вони допомагають орієнтуватися в просторі, і тому ендодерма стебла називається крохмалоноснимпіхвою.Крім того, у різних рослин у первинній корі можуть формуватися вмістилища виділень, можуть зустрічатися кристалоносні клітини, це пластична частина стебла.
Центральний осьовий циліндрвін починається з перециклічної склеренхіми. Перицикл у стеблі перероджується у склеренхіму та частково в паренхіму. Періциклічна склеренхіма може бути на поперечному розрізі у вигляді суцільного кільця або у вигляді окремих тяжів.
За перициклом розташовані провідні пучки, зазвичай колотиральні; рідко біколотиральні оточені клітинами основної паренхіми.
Розташування провідних пучків по-різному у однодольних і дводольних рослин.
- У однодольних розташоване дифузно (розсіяно)
- У дводольних по колу
Таке різне розташування провідних пучків пов'язане з характером об'єднання провідних пучків листя та стебла.
У дводольних провідні пучки листа з'єднуються зі стеблом у своєму вузлі. ![]()
У однодольних листові пучки заходять у центральний циліндр згинаються і з'єднуються зі стебловими пучками, проходячи через 1-2 вузли. 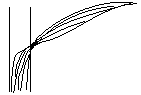
У дводольних з листа в стебло заходить 2-3 листових пучки, а у однодольних у зв'язку з паралельним жилкуванням - безліч.
У однодольних рослин усі провідні пучки закриті, тобто. не мають камбію, а у дводольних відкритіпучки, тобто. є камбій.
Товщина стебла однодольних рослин по всій довжині залишається однаковою, а у дводольних верхня частинастебла трохи менше, ніж у нижній частині.
Стебло. Морфологія та функції стебла.Стебло - подовжена втеча вищих рослин, службовець механічною віссю, також виконує функцію провідної та опорної бази для листя, нирок, квіток. Стебла виконують дві основні функції - несучу та ведучу. Стебла з'єднують усі частини втечі в єдину систему. Завдяки стеблу забезпечується збільшення поверхні рослини шляхом її розгалуження, утворення та вигідніше розташування листя та генеративних органів. Стебло забезпечує висхідний і низхідний рух води та розчинених у ній речовин. Молоді пагони виконують функцію фотосинтезу. Видозмінені стебла виконують ще й додаткові функції, наприклад, накопичення та зберігання поживних речовин у багаторічних стеблах, а також запасання води у спеціалізованих стеблах, зачіпки інших рослин, захист, забезпечення вегетативного розмноження рослин.
Стебла рослин дуже різноманітні та відрізняються за багатьма ознаками. За консистенцією стебла бувають: трав'янистими або дерев'янистими, пустотілими (соломинка) або наповненими (з серцевиною). ), багатогранні (види лободи, щавлю), сплющені (наприклад, тонконіг стиснутий, ситник членистий). За характером зростання та розміщенням у просторі виділяють наступні видистебел: прямостоячі (міцні, переважно ортотропні стебла з добре вираженими міжвузлями, більшість видів вищих рослин) повзучі (переважно плагіотропні стебла вкорінюються у вузлах, з добре вираженими міжвузлями; наприклад, живучка повзуча, суниці повзучий опори за годинниковою стрілкою або навпаки, наприклад, хміль звичайний, берізка польова, повилика, квасоля звичайна) лазячі або чіпкі (стебла з погано розвиненою механічною тканиною зачіплюються за допомогою вусиків; наприклад, огірки, гарбуз звичайний, горох посівний) погано розвиненою механічно тканиною, що стелиться по поверхні ґрунту: наприклад, грижник голий, мшанки лежача, звіробій, що стелиться, стелюшок) висхідні (нижня частина стебла стелиться по поверхні ґрунту, верхня - спрямована вертикально; наприклад чебрець, зірочка, вероніка лікарська) укорочені ( виражене, з дуже зближеними вузлами, короткими міжвузля та зібраним у прикореневу розетку листям, утворює квіткову стрілку; наприклад, подорожник, кульбаба, лілійники, пролісок). Видозміни стебел.
Видозмінене стебло є органом, призначеним для накопичення запасних поживних речовин, які необхідні рослині, щоб пережити період спокою. Крім того, видозмінені стебла нерідко беруть участь у розмноженні та розповсюдженні рослин. Щоб бути віднесеним до цієї групи, видозмінене стебло має мати всі ознаки звичайного стебла, тобто мати характерну для нього будову, що означає наявність власне стебла з верхівковою точкою росту і листям, що розташовуються по спіралі (чергово або супротивно) і несуть у своїх пазухах нирки. . Відмінність видозміненого стебла отнормального полягає в тому, що він не обов'язково зростає над землею, зростання його часто буває більш-менш горизонтальним і зазвичай є вмістищем запасних поживних речовин.
Існує шість основних типів видозмінених стебел: бульба кореневища бульбоцибулина цибулина вус син
Виникнення первинних тканин та первинна анатомічна будова.Анатомічна будова стебла рослин зумовлена його основними функціями. Для нього характерний розвиток механічної та провідної тканин рослин. Крім того, для стебла характерна складна системамеристем - верхівкові, бічні та вставні, що визначають наростання його протягом тривалого часу та виникнення нових органів.
Стебло рослини виникає з апікальної меристеми, з якої диференціюються три шари тканин: покривна, провідна, основна. Спочатку ці тканини представлені так званою первинною меристемою (промеристемою), що складається з протодерми, прокамбію та основної меристеми. Первинні провідні тканини розвиваються з прокамбію. Перші елементи флоеми диференціюються із зовнішніх, розташованих до периферії клітин прокамбію. Первинна флоема представлена тонкостінними недовговічними подовженими клітинами і носить назву протофлоеми, а зовнішні клітини її можуть бути представлені механічними волокнами. внутрішніх клітинпрокамбію і визначаються загалом як протоксилема. Пізніше всередину від протофлоеми диференціюється має більш менш типове для флоеми будову. Зовнішньо від протоксилеми формується метаксилема, що складається з трахеїд або трахей з більш потовщеними здерев'янілими стінками.
Таким чином, за рахунок діяльності прокамбію та решти меристеми апекса виникає первинна будовастебла рослини. У однодольних рослин весь прокамбій диференціюється в елементи первинних провідних тканин. Стебла однодольних, особливо трав'янистих (злаків), в порівнянні зі стеблами дводольних рослин мають простішу будову, для них характерна в основному первинна будова. , обсяг яких збільшується за рахунок поділу клітин камбію. Пучки, що складаються тільки з первинних тканин, як у однодольних, закриті, пучки з камбієм, характерні для дводольних, - відкриті. Пучки, що проводять, закриті, колатеральні, рідше концентричні. З механічних тканин найбільш розвинена склеренхіма, коленхіма зустрічається у небагатьох рослин. Вторинне потовщення у трав'янистих однодольних немає.
Формування тканин стебла вторинної будови
Вторинна будова стебла характерна для однорічних та багаторічних трав'янистих, дерев'яних дводольних, а також голонасінних рослин. У дводольних рослин первинна будова дуже недовговічна, і з початком діяльності камбію утворюється вторинна структура. Залежно від закладки прокамбію формується кілька типів вторинної будови стебла. Якщо тяжі прокамбію розділені широкими рядами паренхіми, то формується пучкова будова, якщо вони зближені так, що зливаються в циліндр - формується непучкова будова.
Рис. 3.24.Пучковий тип будови стебла дводольної рослини: А - конюшина: 1 - епідерма; 2 – хлоренхіму; 3 – склеренхіма перициклічного походження; 4 – флоема; 5 – пучковий камбій; 6 – ксилема; 7 - міжпучковий камбій
Пучкова будова стеблазустрічається у таких рослин, як конюшина, горох, жовтець, кріп (рис. 3.24). У них прокамбіальні тяжі закладаються в одне коло по периферії центрального циліндра. Кожен прокамбіальний тяж перетворюється на колатеральний пучок, що складається з первинної флоеми та первинної ксилеми. Надалі між флоемою та ксилемою з прокамбію закладається камбій, що формує елементи вторинної флоеми та вторинної ксілеми. До периферії органу відкладається флоема, а центру - ксилема, причому ксилемы відкладається більше. Первинні флоема та ксилема залишаються на периферії пучка, а вторинні елементи примикають до камбію. Для стебел дводольних рослин характерно формування відкритих колатеральних або біколатеральних пучків (рис. 17, див. цв. вкл.).
Також для стебел дводольних рослин характерна диференціація первинної кори,до складу якої входять: коленхіма (кутова (рис. 18, див. цв. вкл.) або пластинчаста), хлорофілоносна паренхіма та внутрішній шар– ендодерма. В ендодермі накопичується крохмаль; таке крохмалоносна піхваграє важливу рольу геотропічній реакції стебел. На межі первинної кори в центральному осьовому циліндрі розташовується перициклічна склеренхіма- суцільним кільцем або ділянками у вигляді напівдуг над флоемою. Серцевина стебла виражена та представлена паренхімою. Іноді частина серцевини руйнується із заснуванням порожнини (див. рис. 3.24).
Непучкова будовахарактерно для деревних рослин (липа) (рис. 19, див. цв. вкл.) та багатьох трав (льон). У конусі наростання прокамбіальні тяжи зливаються і утворюють суцільний циліндр, видимий на поперечному зрізіу вигляді кільця. Кільце прокамбію назовні формує кільце первинної флоеми, а всередину - кільце первинної ксілеми, між якими закладається кільце камбію. Клітини камбію діляться (паралельно поверхні органу) і назовні відкладають кільце вторинної флоеми, а всередину - кільце вторинної ксілеми у співвідношенні 1:20. Непучкову будову розглянемо з прикладу багаторічного деревного стебла липи (рис. 3.25).
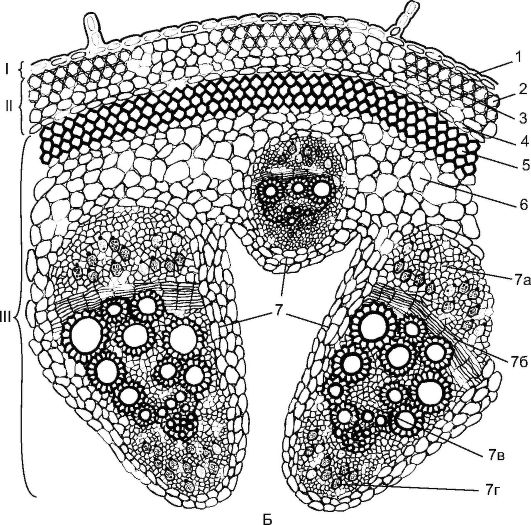
Рис. 3.24.(Продовження) Б - гарбуз: I - покривна тканина; II – первинна кора; III – центральний осьовий циліндр; 1 – епідерма; 2 - кутова коленхіма; 3 – хлоренхіму; 4 – ендодерма; 5 – склеренхіма; 6 – основна паренхіма; 7 - біколатеральний судинно-волокнистий пучок: 7а - флоема; 7б – камбій; 7в – ксилема; 7г - внутрішня флоема
Молодий пагін липи, що утворився навесні з нирки, покритий епідермою. Усі тканини, що лежать до камбію, називають корою. Кора буває первинна та вторинна. Первинна корапредставлена пластинчастою коленхімою, що розташовується відразу під епідермою суцільним кільцем, хлорофілоносною паренхімою та однорядною крохмалоносною піхвою. У цьому шарі знаходяться зерна «крохмалю, що оберігається», які рослина не витрачає. Вважається, що це крохмаль бере участь у підтримці рослиною рівноваги.
Центральний осьовий циліндр у липи починається з перициклічної склеренхіми над ділянками флоеми. Внаслідок діяльності камбію виникає вторинна кора(від камбію до перидерми), представлена вторинною флоемою, серцевинними променями та паренхімою вторинної кори. Кору з липи заготовляють, знімаючи до камбію, особливо легко це робити навесні, коли клітини камбію активно діляться. Раніше кору липи (лико) використовували для плетіння лаптей, виготовлення коробів, мочалок та ін.
Трапецієподібна флоема розділена трикутними первинними серцевинними променями, що пронизують деревину до серцевини. Склад флоеми у липи неоднорідний. У ній є складові твердий луб дерев'яні луб'яні волокна, а м'який луб представлений ситовидними трубками з клітинами-супутницями і луб'яною паренхімою. Луб втрачає здатність проводити органічні речовини зазвичай через рік та оновлюється новими шарами за рахунок діяльності камбію.
Камбій утворює і вторинні серцеві промені, але вони не доходять до серцевини, гублячись у вторинній деревині. Серцеві промені служать для просування води та органічних речовину радиальному напрямку. У паренхімних клітинах серцевинних променів до осені відкладаються запасні поживні речовини(крохмаль, олії), що витрачаються навесні на зростання молодих пагонів.
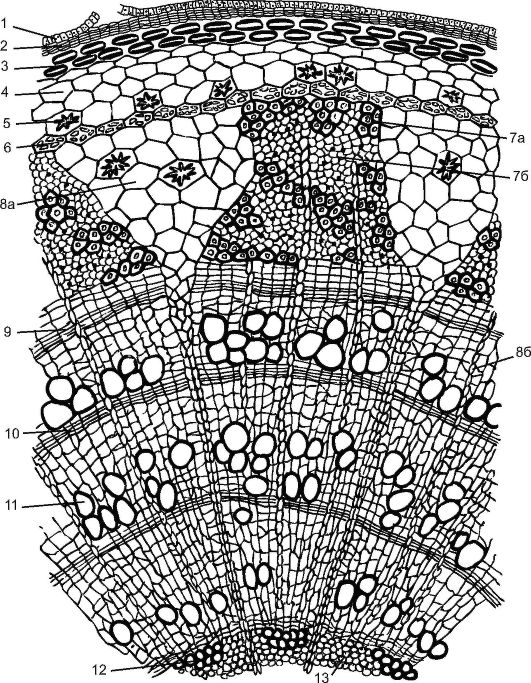
Рис. 3.25.Поперечний розріз трирічної гілки липи: 1 – залишки епідерми; 2 – пробка; 3 - пластинчаста коленхіма; 4 – хлоренхіму; 5 – друзи; 6 – ендодерма; 7 - флоема: 7а - твердий луб (луб'яні волокна); 7б - м'який луб - (ситоподібні трубки з клітинами-супутницями та луб'яна паренхіма); 8а - первинний серцевинний промінь; 8б - вторинний серцевинний промінь; 9 – камбій; 10 – осіння деревина; 11 – весняна деревина; 12 – первинна ксилема; 13 - паренхіма серцевини
Вже влітку під епідермою закладається феллоген та формується вторинна покривна тканина – перидерма. До осені, з утворенням перидерми клітини епідерми відмирають, але їх залишки зберігаються протягом 2-3 років. Нашарування багаторічних перидерм формує кірку.
Шар ксілеми, утворений камбієм, у деревних рослин значно ширший, ніж шар флоеми. Деревина функціонує протягом кількох років. Відмерлі клітини деревини не беруть участь у проведенні речовин, але здатні підтримувати колосальну тяжкість крони рослини.
Склад деревини неоднорідний, до неї входять: трахеїди(рис. 20, див. кол. вкл.), трахеї, дерев'яна паренхімаі лібріформ.Деревина характеризується наявністю річних кілець.Ранньою весною, коли в рослині виникає активний рух соку, камбій у ксилемі формує широкопросвітні та тонкостінні провідні елементи - судини та трахеїди, а з наближенням осені, коли ці процеси завмирають і діяльність камбію слабшає, виникають вузькопросвітні товстостінні судини, товстостінні судини, трасеїни. Таким чином, утворюється річний приріст, або річне кільце (від однієї весни до іншої), добре помітне на поперечному зрізі. За річними кільцями можна визначити вік рослини (див. рис. 3.25).
Особливості будови стебла дводольних:
1) зростання стебла у товщину (за рахунок діяльності камбію);
2) добре диференційована первинна кора (коленхіма, хлорофілоносна паренхіма, крохмалоносна ендодерма);
3) біколатеральні та колатеральні пучки тільки відкритого типу(З камбієм);
4) судинно-волокнисті пучки розташовані по кільцю або зливаються (непучкова будова);
5) наявність серцевини;
6) для деревних рослин характерна наявність у ксилемі річних кілець.
Особливості будови кореневищ дводольних.Покривною тканиною дводольних кореневищ може бути епідерма, а у багаторічних кореневищ епідерма змінюється перидермою. Первинна кора представлена паренхімою і ендодермою з плямами Каспарі. При цьому ширина первинної кори наближається до ширини центрального циліндра. Будова центрального осьового циліндра, судинно-волокнистих пучків та його розташування у ньому мають самі особливості, що й надземних стебел.
Залежно від екологічних умов, у яких формується той чи інший вид рослини, створюються різні особливостіструктури його органів, зокрема й стебла.
Крім того, стебло у ряді випадків виконує функції інших органів. І тут зміни структури стебла особливо суттєві і зазвичай зараховуються до розряду аномалій. Своєрідною будовою стебла характеризуються ліани. Особливо багато представлені ліани в тропіках, де вони найрізноманітніші і потужно розвинені. Але й у помірних широтах росте досить ліан. Хміль, ломонос, плющ, виноградна лоза, горох, квасоля, гліцинія, берізка та ін - справжні ліани, не настільки розкішні, як у тропіках, але все ж таки мають у структурі стебла риси, властиві всім ліанам.
Найбільш характерною рисою структури стебла ліан є досить різко виражена розчленованість провідної системи та сильний розвитокпаренхімних серцевих променів. Навіть у ліан з прекрасно вираженим деревним стеблом (виноградна лоза) серцевинні промені складаються з паренхіми, що мало відрізняється від паренхіми кори і серцевини, причому ці серцевинні промені дуже високі, тягнуться на великій відстанівздовж довжини стебла, нагадуючи довгі, вертикально поставлені вздовж стебла стрічки, що розсікають масив центрального циліндра на окремі сектори. У типових деревних рослин серцевинні промені внаслідок незначності їхньої висоти на тангентальних розрізах деревини мають вигляд коротких веретеноподібних скупчень клітин. Ще різкіше розчленовування центрального циліндра стебла виявляється у тропічних ліан. У деяких тропічних ліан розкидані у різних місцях деревини центрального циліндра паренхімні клітини починають посилено ділитися та розростатися, розсовуючи окремі ділянки судинної системи. Внаслідок цього старі стебла таких ліан відрізняються дуже неправильною та вибагливою будовою. Загалом будова стебла деревних ліан нагадує будову стебла деяких трав'янистих рослин.
У ряду рослин, що виростають у пустельних місцевостях Африки та Америки, де взагалі дощі випадають дуже рідко і те протягом короткої пори року, формі та структурі стебла властиві особливі специфічні риси. У таких рослин стебла м'ясисті, зелені, приймають. різну форму- кулі, пластинки або забезпечені широкими пластинчастими ребрами. Такі стебла виконують функцію листя. Листя часто дуже дрібне і іноді має вигляд сухих колючок. Рослини з м'ясистими стеблами та редукованим листям називають стебловими суккулентами. До них відносяться різні видиопунцій, кактусів, молочаїв.
Редукція листових платівокдо сухих колючок веде до значного зменшення поверхні, що випаровує всієї рослини, що складається у своїй надземній частині цілком зі своєрідно видозміненого стебла. Саме стебло перетворюється на орган, пристосований для накопичення запасів води. Це дозволяє рослині запасати вологу в дощові періоди і повільно витрачати її в пору бездощів.
У будові покривних тканин більшості типів суккулентних стебел є пристосування до можливого зниження втрати рослиною води через транспірацію. При цьому має бути сильно редукована і система провідних тканин. Механічні тканини всередині суккулентних стебел стають непотрібними та майже відсутні.
Клітинний сік клітин тканин суккулентних стебел складається з розчинів різних кислот, цукрів та слизових речовин. Ці речовини сприяють зниженню віддачі води рослиною. Тканина суккулентних стебел дуже однорідна і складається переважно з тонкостінної паренхіми, пронизаної слаборозвиненими судинними пучками: судин у кожному пучку небагато, всі вони вузькопорожнинні та примітивної структури. Слабкий розвиток судинних пучків цілком відповідає незначному розвитку листя, тому що лист у своєму розвитку стимулює утворення судинно-волокнистих пучків стебла. У типових, добре виражених суккулентних стеблах вторинної ксилеми утворюється мало внаслідок слабкої камбіальної діяльності, судинна системаскладається переважно з первинних елементів, що виникли у васкулярній меристемі. Отже, і фізіологічно, і з погляду історії розвитку формування слабкої провідної системи в стеблах суккулентних рослин цілком зрозуміле.
Природно, що серед сукулентів немає деревних форм, усі вони за своєю структурою – трав'янисті рослини.
У рослин так званого склеренхімного типу, що теж виростають у посушливих місцях, розвинені нормальні листя і стебла, провідна система складається з потужно розвинених судинно-волокнистих пучків або продуктів їх злиття. Судинні елементи в таких пучках широкосмугові, численні і високо організовані, ніж у сукулентів. Побічні елементи у деревних рослин цієї групи виражені чітко, а стеблах трав'янистих рослин різко виражено здеревнення оболонок клітин основний паренхіми. Механічна система розвинена сильно, анатомічні елементи її товстостінні.
Стебла рослин, що ростуть у воді, - так званих водних рослин - при відносно багатому розмаїтті форм мають один загальний структурною ознакою: у них добре розвинена система міжклітинних ходів та міжклітинників. Міжклітини у вигляді більш-менш великих порожнин утворюють систему повітряних ходів, що пронизують як серцевину, так і кору стебла. При цьому особливо сильно розвинена серцевина, що займає значну частку загального діаметра стебла. У водних рослин, особливо серед представників класу однодольних, механічна системаіноді переважає над провідною системою. Групи судинних анатомічних елементів, зібраних у пучки, зазвичай бувають порівняно незначних розмірівмасиви ж, що складаються з механічних елементів, зосереджені переважно по периферії стебла, досягають порівняно з судинними пучками великої потужності.
Ще різкіше відхиляється від нормального типубудова підземних стебел, що виконують функцію вмістищ запасів, - кореневищ та бульб. Кореневище є багаторічною втечею і зазвичай досягає значно більшої товщини, ніж надземні пагони, де б вони не виростали, - у воді чи іншому середовищі. У зв'язку з основним призначенням кореневища - служити сховищем запасних речовин - паренхіматизація його тканин є найбільш характерну рисуструктури. Запасаючою тканиною в типових кореневищах служить серцевина, що складається з округлих паренхімних клітин, тонкостінних, з невеликими міжклітинниками між ними. За своєю масивністю серцевина у кореневищах переважає над іншими тканинами. Механічні та провідні тканини, поступаючись за потужністю серцевині та корі, у кореневищі виражені слабше, ніж у надземних пагонах.
Структура кореневища як багаторічної втечі складається під впливом різноманітних формують факторів зовнішнього середовища. Вплив цих чинників може бути як безпосереднім, і діючим через кореляцію у розвитку органів рослини: так, ступінь розвитку листя, утворення квіткових пагонів, відкладання запасних речовин впливають структуру кореневища. Особливо помітна залежність будови кореневища від рівня розвитку на них листя.
Під час підземного зростання, коли на кореневищі знаходяться лише лускоподібні дрібні листочки, відповідно формується дуже слабка провідна система, що в ксилемній частині складається з рідкісних вузькопорожнинних судин примітивної структури. Після того як почне розвиватися надземна плодоносна втеча з нормальним великим листям, посилюється активність камбію і внаслідок цього розвивається деревина, а також збільшується число і просвіт судин, які мають більш досконалу структуру в порівнянні з утвореними до початку зростання надземної втечі.
Своєрідною морфологічною особливістюКореневища дуже короткі міжвузля. Очевидно, незначність довжини міжвузлів перебуває у тісній кореляції зі слабкою диференціацією листя на кореневище.
На кореневище за своєю морфологією в деяких відносинах схоже інше видозміна підземної втечі - бульба. Основна відмінність між кореневищем і бульбою полягає в тривалості існування: кореневище - багаторічна втеча, бульба зазвичай існує один-два роки. Тому в клубні виявлено лише ті структурні рисиякі кореневища мають на першій стадії формування своїх тканин. Переважна більшість паренхіми в бульбі ще виразніше, ніж у кореневищі.
Ступенем паренхіматизації тканина бульби нагадує тканину стебла сукулентних рослин. Так само, як у стеблах суккулентних рослин, вся маса тканини бульби за винятком покривної (перидерми) є тканиною, що запасає, пронизану слаборозвиненими і примітивно побудованими пучками провідної системи. Поряд з редукцією провідної тканини в нормальному бульбі зазвичай зовсім не утворюється механічна тканина. Тканини, що складають бульбу, одноманітно побудовані та пристосовані до виконання лише однієї функції – відкладення запасів.

Типовим прикладом структури бульби може бути бульба картоплі. На малюнку зображено фрагмент поперечного розрізу бульби одного з сортів картоплі («Вольтман»), із центральної частини його. Для зручності демонстрації тканинної структури крохмаль, удосталь заповнює всі клітини, не зображений. Паренхіма бульби лише одному місці розсікається вузькою смужкою дрібніших клітин, що з чотирьох груп. У кожній групі знаходиться 1-3 вузькопорожнинні судини, решта тканини складається з меристемоподібних клітин, серед яких знаходиться кілька ситоподібних трубок. Судини, що відрізняються слаборозвиненою структурою, примітивні, причому деякі з них перебувають у стані облітерації та руйнування.
Така тканина, що складається з примітивних і слаборозвинених судинних елементів, розташована кільцем по всьому бульбі, являючи собою центральний циліндр. Назовні розташована первинна кора, усередині - серцевина. У бульбах, що мають лише лускоподібні подоби листя, вторинних тканин внаслідок діяльності камбію не утворюється.
Якщо ви знайшли помилку, будь ласка, виділіть фрагмент тексту та натисніть Ctrl+Enter.


