Тип судини на поперечному зрізі стебла кукурудзи. Первинна будова стебла
(Комплекси: "Ксилема", "Флоема". Видільні тканини)
Провідні тканини забезпечують пересування речовин у рослині. Висхідний струм (водні розчини мінеральних речовин) від коренів до надземних органів проходить судинами і трахеїдами. Східний струм (розчин органічних речовин) рухається ситоподібними трубками флоеми (лубу) від листя в стебла і коріння. Клітини провідних тканин у тілі рослин входять до складу спеціальних комплексів. Трахеї та трахеїди є частиною ксилеми, ситоподібні трубки з клітинами-супутницями - частина флоеми. До складу ксилеми та флоеми, крім провідних тканин, входять клітини склеренхіми та паренхіми. Ксилема та флоема утворюються з прокамбію та камбію.
У той час як гриб може викликати руйнівні хвороби листя листя та стебел, мало відомо про його здатність заражати коріння. Ми спостерігали структури, створені іншими кореневими патогенними грибами, включаючи гінекологічні гіфи, гіфоподії та мікросклеротиа. Мозаїчна картина інфекції обумовлена тим, що окремі епідермальні та кортикальні клітини заражаються міжклітинними гіфами, а оточуючі клітини неінфіковані, картина відрізняється від описаної для листя.
Цікаво відзначити, що конідії з копитами, зазвичай обмежені ацервулами, також були виявлені, заповнюючи епідермальні клітини та кореневі волоски. Двадцять вісім відсотків рослин, заражених ґрунтовим інокулятом, заразилися у надземних частинах рослин, що вказує на те, що коренева інфекція може призвести до безсимптомної системної колонізації рослин. Ці спостереження свідчать, що коренева інфекція може бути важливим компонентомциклу хвороби антраклози кукурудзи Хвороба може вражати більшість рослинних тканин, хоча найімовірнішими є руйнівні форми хвороби цвілі та розсади.
Робота 22 Гарбузи звичайної, що ведуть тканини в стеблі - Cucúrbita pépo L .
Судини (трахеї) - це трубки, що утворюються з вертикального ряду клітин прокамбію або камбію, у яких товщають і одревесневають поздовжні стінки з облямованими порами, а в поперечних стінках утворюються одна або кілька перфорацій і відмирає вміст. За характером потовщення стінки розрізняють судини: кільчасті, спіральні, сходові, сітчасто-пористі та ін.
Протягом останніх кількох десятиліть він став визнаним одним з переважаючих патогенних мікроорганізмів кукурудзи Північної Америки. Під час форми хвороби листя, видимі ураження формуються спочатку на нижньому листі і поступово просуваються вгору в міру дорослішання рослини. Значна частина наших знань про кукурудзяну антракнозу походить від вивчення фази хвороби листя хвороби. З ультраструктурних досліджень відомо, що після утворення апресора утворюється штифт, що проникає, який проникає в епідермальну клітину через кутикулу.
Трахеїди на відміну судин не багатоклітинні трубки, а прозенхімні клітини. У стінах їх є облямовані пори. З трахеїди трахеїду вода піднімається повільніше, ніж у посудині, т.к. просочується крізь пори. Стінки трахеїд та судин у молодому віціцелюлозні. Потім вони з'являються вторинні потовщення, які мають вигляд спіралі, кілець тощо. Такі потовщення просочуються лігніном (одревесневають) і надають стінкам міцність.
Усередині епідермальної клітини утворюється неаргулярна первинна гіфа. великого діаметру. Гіфа росте біотрофно і може утворювати одну або декілька гілок, які поширюються на сусідні епідермальні або мезофільні клітини. Гриб росте між рослинною плазматичною мембраноюі стінкою клітини рослини і поширюється від клітини до клітини таким чином протягом 36-48 годин. До 48 до 72 год відбувається перехід на некротрофічне зростання, яке типове для виробництва великої кількостігіф меншого діаметра, які широко колонізують між- та внутрішньоклітинні простори тканини, викликаючи загибель клітин-господарів до колонізації.
Ситоподібні трубки складаються з подовжених живих клітин - члеників, що розташовані одна над одною. Поперечні стінки члеників ситоподібних трубок є ситоподібними пластинками. Вони пронизані безліччю дрібних отворів (перфорацій), що утворюють канальці. Через ці отвори з клітини в клітину йдуть тяжі цитоплазми. Ядер у цих клітинах немає. Поруч із члеником ситовидної трубки розташовується одна або кілька клітин-супутницьз густою цитоплазмою та ядром.
Після початку тканинного підсмажування, вузькі вторинні гіфи від первинних гіф у клітинах мертвого господаря та випромінюють у навколишні тканини. Отже, патоген, мабуть, здатний приймати різні способи життя, залежно від цього, який господар він колонізує. Таким чином, здатність викликати захворювання та здатність колонізувати тканину господаря, мабуть, є окремими рисамиіз незалежним генетичним контролем. Фактично, багато грибів, які зазвичай вважаються причинами збудника листяних хвороб, також можуть заражати своїх господарів, вторгаючись до коріння.
Порядок роботи:
Постійний препарат поздовжнього зрізу стебла гарбуза звичайного - Cucúrbita pépo L. – розглядають при малому збільшенні. Знаходять ділянку зрізу з судинами та переходять на велике збільшення.
При великому збільшенні вивчають кільчасті, спіральні, сітчасті, пористі та сітчасто-пористі судини. Стінки кільчастих та спіральних судин целюлозні, неодревеснілі. На препараті вони блакитного кольоруз добре помітними здерев'янілими потовщеннями червоного кольору. Стінки судин великого діаметра, що здерев'яніли. Вони забарвлені у червоний колір. Окремі світлі ділянкиу них – пори. Це сітчасто-пористі судини.
Однак, кореневі інфекції, що супроводжуються системною колонізацією рослини, не були показані, і жодні дослідження не описували, як гриб може приєднуватися, проникати та колонізувати коріння рослин. Такі знання були б корисними для розуміння розвитку хвороби та розробки стратегій боротьби з хворобами. Грибкові штами та умови культури. Культури підтримували при 25 ° С на середовищі агару з картопляною декстрозою з безперервним освітленням від флуоресцентного джерела світла.
Протопласти були звільнені після 4-5 годин травлення. Аналіз стабільності трансформантів та патогенності. Після інкубації протягом 24 годин вибирали одну колонію. Цей процес повторювався один раз. Трансформанти аналізували на морфологію колоній, швидкість зростання та вірулентність.
Замальовують судини різних типів, деревини стіни розфарбовують червоним кольором, роблять відповідні позначення (рис. 21).
Мал. 21. Типи судин.
На тому препараті при малому збільшенні знаходять ситоподібні трубки з клітинами-супутницями.
Переходять на велике збільшення та вивчають будову ситоподібних трубок, ситоподібних пластинок та клітин супутниць. У ситоподібних платівках видно дрібні отвори. Вузькі клітини-супутниці мають великі ядра та густу цитоплазму.
Перед посівом все насіння обробляли, покриваючи його фунгіцидом трифлоксистробіном і дозволяючи їм сушити при кімнатній температуріпротягом 24 годин. Насіння промивали в 70% етанолі протягом 5 хв і одразу промивали водою для видалення фунгіциду перед інокулюванням. Використовувалися три методи інокуляції коріння. Для вивчення ранньої колонізації коріння ми інокулювали коріння кукурудзи з насіння, яке було проростане в чашках Петрі. Насіння поміщали в чашки Петрі в наборах по чотири на мокрому фільтрувальному автоклавному папері і кожні 2 дні поливали розчином добрива 5 г літра -1 Пітера.
Замальовують кілька члеників ситовидної трубки з тяжами цитоплазми, однією-двома ситовидними пластинками і клітинами супутницями, роблять позначення (рис. 22).
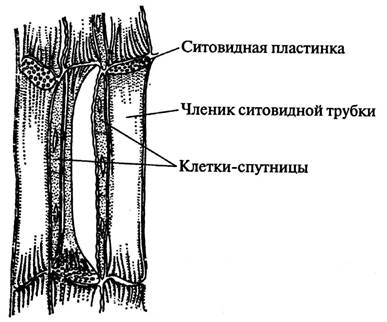
Мал. 22. Елементи флоеми на поздовжньому зрізі
Провідні пучки
В органах рослин ксилема та флоема розташовуються спільно, утворюючи провідні пучки. Якщо між ксилемою та флоемою є камбій - пучки відкриті, якщо камбію немає - закриті. Відкриті пучки характерні для дводольних покритонасінних і голонасінних рослин, закриті - для однодольних покритонасінних і папоротьподібних. До складу пучків нерідко входить механічна тканина – склеренхіма. Такі пучки називають судинно-волокнистими.
Саджанці інокулювали 3-денним післягермуванням з використанням методу кореневого вимочування, або агару шви. Конідії фільтрували через три шари марлі і потім промивали двома раундами ресуспендування в дистильованій стерильній воді з подальшим центрифугуванням.
Коріння вимочували протягом 1 години в суспензії 107 конідій мл -1. безпосередньо на відкритому корінні розсади. Після інокуляції саджанці спостерігалися щодня під мікроскопом, починаючи з 1 постінокуляції. Ще 5 мл вермікуліту поміщали поверх агарових пробок, додавали одне зерно кукурудзи та додавали ще 5 мл вермікуліту для створення близько 1 см вермикулітового покриття. Десять розсади було підготовлено для кожного випробування. Коли проростки проростали, парафілм видаляли, а саджанці поливали через день, використовуючи добрива 5 г літра -1 Пітера.
Ксилема та флоема в закритих пучках за походженням первинні, т.к. формуються з прокамбія. У відкритих пучках значна частинаксилеми та флоеми виникає в результаті діяльності камбія, тобто. вони вторинніпоходження, тому у складі відкритих пучків є первинна та вторинна ксилема, а також первинна та вторинна флоема. За взаємним розташуванням ксилеми і флоеми, що проводять пучки поділяються на колатеральні, біколатеральні, концентричніі радіальні(Рис. 23).
Експеримент проводився чотири рази. Для експериментів, що вимагають рослин понад 3 тижні, саджанці спочатку вирощували у вермикуліті та інокулювали агаровими пробками, як описано вище. Рослини переносили в 3 або 10-літрові горщики, що містять суміш 40% торфу, 40% суглинку, 10% вермикуліту та 10% компосту. Інтенсивність світла у теплиці протягом дня становила від 120 до 200 мікронних хв м-2.
Рослини, які використовуються в тепличних експериментах, щодня зрошувалися водою і двічі на тиждень за допомогою добрива Пітера. Від листя, стебел і коріння розрізали секції діаметром від 2 до 3 мм. Зразки інкубували при 25 °С протягом 10 днів і щодня аналізували за допомогою світло-польової та флуоресцентної мікроскопії. Цей експеримент повторювали три рази, причому кожна реплікація складалася з семи щеплених рослин та семи щеплених рослин. Обробка тканин та мікроскопія.
Мал. 23. Схема будови різних типів провідних пучків:
1 – колатеральний закритий; 2 – колатеральний відкритий; 3 – біколатеральний відкритий; 4 – амфівазальний концентричний закритий; 5 – амфікрибральний концентричний закритий; 6 – радіальний тетрархний пучок
Робота 23 Закритий колатеральний пучок на поперечному зрізістебла кукурудзи звичайної - Zéa máys L .
Бари = 100 мкм, 20 мкм та 10 мкм. Усі фотографії були отримані за допомогою флуоресцентного мікроскопа, якщо не вказано інше. Гіфи подовжуються в конідієносці, які утворюють зрілі первинні конідії. Зверніть увагу на незрілі конідієносці. Розглядається з флуоресценцією та освітленням яскравого поля. Щетинки можуть утворюватися усередині клітини. Поперечний переріз кореня кукурудзи; 14 пікселів на дюйм. Коркові клітини колонізуються гіфами та овальними конідіями. Коренева епідермальна клітина, заповнена конідія фальцату.
Фарбування коричневого кольоруможе спостерігатися у інфікованого коріння, але не спостерігається ніякого некрозу. Гіфи колонізують клітини кори всередині та внутрішньоклітинно. Поперечний переріз кореня; 14 пікселів на дюйм. Міжклітинні товсті меланізовані гіфи, що колонізують кору кореня. Колонізація кореневої кори.
Порядок роботи:
При малому збільшенні постійному препараті поперечного зрізу стебла кукурудзи вибирають найбільший пучок (ближче до центру зрізу).
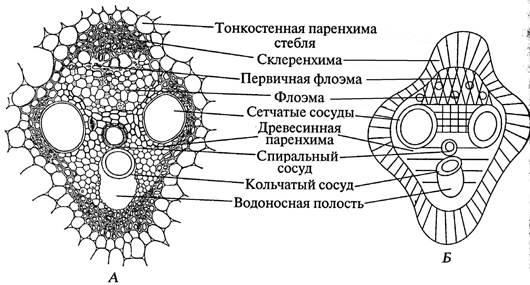
За великого збільшення розглядають пучок. Зазначають, що ксилема та флоема в ньому оточені склеренхімою – судинно-волокнистий пучок. На препараті елементи ксилеми і склеренхіму, що здерев'яніли, пофарбовані в червоний колір. Під склеренхімою, примикаючи один до одного, розташовуються первинна ксилема (навернена до центру стебла) і первинна флоема (навернена до периферії стебла). У ксілемі добре помітні червоні судини, що здервіли, а також водоносна порожнина. Усього на зрізі видно три-п'ять судин, дві з них найбільші. Це пористі судини.
Гіфи сильно колонізують епідерміс та кору та досягають ентодерми. Гіфи колонізують ентодермію, флоему та ксилему. Поперечний переріз кореня; 28 пікселів на дюйм. Перетин неінфікованого 16-денного кореня кукурудзи. Жовту аутофлуоресценцію можна спостерігати в епідермісі кукурудзи, ендодермісі, флоемі та ксилемі. Бари = 10 мкм, 20 мкм та 100 мкм. Бари = 10 мкм та 20 мкм. Фотографії були отримані з використанням мікроскопії з яскравим полем, флуоресцентної мікроскопії або під стереомікроскопом.
Колонізація третьої листової оболонки з коренеплідної рослини; 28 пікселів на дюйм. Гіфи ростуть усередині листового трихому. П'ятимікрометровий поперечний переріз листа кукурудзи із посівної рослини; 28 пікселів на дюйм. Від 1 до 28 т/д інфікованого коріння контролювали колонізацію грибів. Для кожного дослідженого моменту часу з кожного з чотирьох незалежних експериментів аналізували щонайменше 20 інокульованих та 5 монокристалічних коренів. Зразки тканини поміщали на предметну скобу мікроскопа, занурювали у краплю води, покривали скляним покривним склом та візуалізували, як описано вище.
У пофарбованій блакитним кольоромфлоемі видно ситоподібні трубки та клітини супутниці. Іноді можна побачити ситоподібні платівки світло-жовтого кольору. Камбію між ксилемою і флоемою немає, отже, пучок, що проводить, закритий.
Схематично замальовують пучок, що проводить (рис. 24).
На цій стадії флуоресценція зазвичай не спостерігалася, тому що накопичення меланіну блокувало світло, хоча менш зрілі флуоресцентні гіфи можна було знайти у зв'язку з меланізованими гіфами. Гіфи широко колонізували поверхню кореня, утворюючи мережу навколо кореня. При 3 т/д гіфах також спостерігалися вторгнення в міжклітинні простори клітин епідермального коріння, хоча на коренях не було виявлено жодних видимих симптомів захворювання. Усередині кореня гіфи роздмухувалися і поширювалися всередині клітини в цитозолі клітин через певні точки дотику з іншими епідермальними і кірковими клітинами.
Мал. 24. Поперечний зріз закритого колатерального пучка стебла кукурудзи. Zéa máys L . А – малюнок, Б – схема.
Робота 24 Відкритий біколатеральний пучок на поперечному зрізі стебла гарбуза звичайного - Cucúrbita pépo L .
Порядок роботи:
На постійному препараті розглядають спочатку при малому, потім при великому збільшення один проводячий пучок. Флоема в пучку знаходиться з двох сторін - зовнішньої та внутрішньої. Під зовнішньою флоемою розташовані ряди тонкостінних живих клітин камбію. Під камбієм добре видно вторинну ксилему з великими судинами, під нею - первинну ксилему з дрібнішими судинами. Описаний провідний пучок називають біколатеральнимчерез наявність двох ділянок флоеми, що примикають з двох сторін до ксилеми. Зовні пучок оточений клітинами основної паренхіми.
Гіфи сильно стиснулися, коли вони проходили через клітинну стінку. Гіфи також можна спостерігати, що ростуть внутрішньоклітинно серед епідермальних та кортикальних клітин. Заражені епідермальні та кортикальні клітини часто заповнювалися гіфами. Гіфи проліферували всередині клітини, заповнюючи більшу частинувнутрішньоклітинного простору і зрештою ставали меланізованими. Гіфи, зважаючи на все, обмежувалися зараженою клітиною, поки внутрішньоклітинний простір не був повністю заповнений гіфами. Тільки деякі епідермальні та кортикальні клітини, як виявилося, колонізувалися таким чином, тоді як оточуючі клітини залишалися неінфікованими, що призводило до мозаїчного картування колонізації рослинних клітин.
Схематично замальовують відкритий біколатеральний пучок (рис. 25).
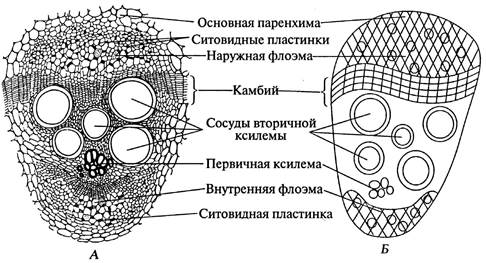
Мал. 25. Відкритий біколатеральний провідний пучок на поперечному зрізі стебла гарбуза звичайного - Cucúrbita pépo L . :А - малюнок; Б - схема
Робота 25 Концентричні пучки на поперечних зрізах кореневищ конвалії травневого ( Convallária majális L . ) та папороті-орляка ( Pterídium aquilínum(L.) Kuhn ex Decken)
Повідомляється, що багато кореневих патогенів інфікують своїх господарів через певні області коренів. Гіфи можна побачити, колонізуючи поверхні зрілих коренів, кореневих капсул, зон подовження коренів та кореневих волосків, що вказує на відсутність переваг кореневої інфекції. Пізніше ці структури розширилися, що призвело до механічного розриву кутикули та утворення мас конусів солоду та характерних щетинок. Цікаво, що конідії сокола утворилися в акербулі, як тільки 5 т/д на поверхнях коренів, але також були виявлені накладки епідермальних клітин та кореневих волосків з роздільною здатністю 10 точок на дюйм.
Порядок роботи:
На постійному препараті поперечного зрізу кореневища конвалії при малому збільшенні знаходять провідний пучок, в якому пофарбовані в червоний колір тканини первинної ксилеми оточують у центрі пучка первинну флоему. Такий пучок називають концентричним амфівазальним.
Переводять мікроскоп на велике збільшення і переконуються без камбію - пучок закритий.
Схематично малюють концентричний амфівазальний пучок.
На постійному препараті поперечного зрізу кореневища папороті - орляка при малому збільшенні знаходять пучок, що проводить, в якому забарвлені в червоний колір тканини первинної ксілеми розташовані в центрі пучка; вони оточені первинною флоемою. Такий пучок – концентричний амфікрибральний.
Переводять мікроскоп на велике збільшення і переконуються у відсутності камбію.
Схематично малюють концентричний амфікрибральний пучок (рис. 26).
Мал. 26.Концентричні амфівазальний (А – малюнок, Б – схема) та амфікрибральний (Г – малюнок, В – схема) пучки на поперечних зрізах кореневищ конвалії травневого - Convallária majális L. (А, Б) та папороті-орляка - Pterídium aquilínum(L.) Kuhn ex Decken (В, Г)
Питання для самоконтролю та підготовки до колоквіуму на теми модульної одиниці 2 "Тканини вищих рослин"
1. Що таке тканина?
2. Якими є відмінності освітніх тканин від постійних?
3. Які тканини називають постійними?
4. Які ознаки покладено основою класифікації постійних тканин?
Освітній тканині
1. Дайте визначення освітньої тканини.
2. Якими особливостями характеризуються клітини освітніх тканин?
3. Який тип поділу характерний для клітин освітніх тканин?
4. Якими є принципи класифікації освітніх тканин?
5. Які шляхи виникнення вторинних меристем?
6. Які меристеми забезпечують наростання органів рослини завдовжки? Завтовшки?
7. Яка роль вставкових меристем?
8. Наведіть приклади виникнення ранових меристем.
9 Що таке калюс?
Постійні тканини
Покривні тканини
1. Які первинні та вторинні покривні тканини вам відомі?
2. Як влаштовано устячки?
3. Чим відрізняються замикаючі клітини продихів від інших епідермальних клітин?
4. Які функції виконує продих? Як замикаючі клітини регулюють розмір гирлової щілини?
5. Які особливості епідерми посухостійких рослин?
6. До складу яких покривних комплексів входить фелема?
7. Із чого і як формується перидерма? Корка?
8. Дайте порівняльну цитологічну характеристику клітин епідерми та фелеми.
9. Чому береста берези бородавчастої білого кольору?
10. Які органи покриті епідермою? Перідермою? Шкіркою?
11. У якому віці стебла багаторічних дерев'янистих рослин покриваються перидермою? Шкіркою?
12Чим покриті. Технічно стиглі коренеплоди та бульби?
13. За якими ознаками можна виділити перидерму на поперечному зрізі стебла дводольної рослини?
14. Як людина використовує покривні тканини рослин?
Основні тканини
1. Чому основні тканини отримали таку назву?
2. Які основні тканини ви знаєте?
3. Які види паренхіми, що асимілює, вам відомі? У яких органах розвивається паренхіма, що запасає? Які запасні речовини відкладаються у її клітинах?
5. Які органи рослин мають поглинаючу паренхіму?
6.Які особливості повітроносної тканини? Для яких рослин вона є характерною?
7.В якій основній тканині закладається феллоген?
Механічні тканини
1. Які види механічних тканин ви знаєте?
2. Яка роль механічних тканин у рослині?
3. Як класифікується коленхіма залежно від характеру потовщення клітинних стінок?
4. Яку функцію, окрім механічної, виконує коленхіма?
5. Чим відрізняються клітини коленхіми від клітин склеренхіми на поперечному та поздовжньому зрізах?
6. Які особливості склереїд (форма клітин, потовщення клітинних стінок, порові канали)?
7. Де і як використовується склеренхіма людиною?
8. У чому цінність склеренхіми льону як прядильної сировини?
Провідні тканини
Ксилема
1. Які функції виконує ксилема?
2. Які гістологічні елементи входять до складу ксилеми?
3. Як відрізняється ксилема за походженням? З яких тканин вона утворюється?
4. У чому різниця між судинами та трахеїдами?
5. Які типи поровності судин та трахеїд вам відомі?
6. У якому напрямку йшла еволюція морфологічних ознаксудин та трахеїд?
7. У яких рослин у ксилемі відсутні судини?
8. Як довго функціонують провідні елементи ксилеми?
9. Які зміни відбуваються в судинах та трахеїдах із припиненням їх діяльності?
10. За рахунок чого відбувається закупорка судин та трахеїд?
11. Що таке тілла?
Флоема
1. Які функції виконує флоема?
2. Які гістологічні елементи входять до складу флоеми?
3. Який онтогенез ситоподібних трубок? Як довго вони працюють?
4. Які особливості протопластів члеників ситоподібних трубок?
5. Що таке первинна та вторинна флоема? З яких тканин вони утворюються?
6. Які рослини у флоемі не мають клітини-супутники?
7. У чому подібність та відмінність гістологічного складу ксилеми та флоеми?
Судинно-волокнисті провідні пучки
1. Що таке пучок, що проводить?
2. Які рослини зустрічаються відкриті пучки? Які закриті?
3. Що таке судинно-волокнистий пучок?
4. Що таке листовий слід?
5. Як називають пучок, що проводить, у листі?
6. Перерахуйте послідовність розташування комплексів тканин у відкритому колатеральному пучку?
7. Судинно-волокнисті провідні пучки яких рослин використовуються людиною?
До категорії: Анатомія рослин
Тканини
Тіло вищих рослин складається з різних клітин, спеціалізованих на виконанні різних функцій. Фізіологічні відправлення клітин визначають їхню структуру. Комплекси клітин, подібних до функцій, а здебільшогоі за будовою, що мають однакове походження та певну локалізацію в тілі рослини, називають тканинами.
Розподіл тканин у органах рослин та його структура тісно пов'язані з виконанням ними певних фізіологічних функцій.
Тканини, що складаються з живих тонкостінних клітин, що інтенсивно діляться, називаються освітніми або меристемами. Верхівкові (апікальні) меристеми розташовані на верхівках стебел і в кінці коренів. Вони зумовлюють зростання цих органів у довжину.
Розгляд апікальних меристем втечі не входить до завдань цього курсу.
До бічних меристем відносяться камбій і фелло-ген. Камбій забезпечує потовщення стебла та кореня. Феллоген утворює корок.
Тканини, що виникають в результаті росту та диференціації клітин - похідних меристем, називають постійними. Поширена в даний час класифікація постійних тканин заснована на їх анатомо-фізіологічних особливостях.
Покривні тканини захищають внутрішні тканинирослин від прямого впливу зовнішнього середовища, регулюють випаровування та газообмін. До них відносяться епідерміс і пробка, що складаються із щільно з'єднаних клітин. У корінні однодольного і молодого коріння деяких дводольних рослин функцію покривної тканини виконує екзодерма.
Механічні тканини зумовлюють міцність рослини. Стіни клітин, що складають ці тканини, потовщені. До механічних тканин відносяться коленхіма, що складається з паренхімних або кілька подовжених клітин з нерівномірно потовщеними целюлозними стінками, і склеренхіма, клітини якої мають рівномірно потовщені здерев'яні стінки. Склеренхіма може бути представлена волокнами та склереїдами ізодіаметричної (кам'янисті клітини), гіллястої та зірчастої форм.
Провідні тканини забезпечують проведення води, ґрунтових розчинів та продуктів асиміляції, що виробляються листям. Тканина, що проводить воду, називають ксилемою або деревиною (особливо у деревних рослин), а тканину, що проводить органічні речовини, утворені рослиною в процесі фотосинтезу - флоемою або лубом. До складу ксилеми і флоеми поряд з власне провідними елементами здебільшого входять механічні та запасні клітини, тому ці тканини називають складними. Провідні тканини за походженням можуть бути первинними та вторинними. Первинні виникають з прокамбію - меристематичної тканини, що закладається на ранніх етапахонтогенезу рослини. Вторинні тканини утворюють камбій, що диференціюється з прокамбію.
Зазвичай флоема і ксилема розташовуються поруч, складаючи пучок, що проводить. Залежно від взаємного розташуванняКсілеми та флоеми розрізняють кілька типів пучків.
Колатеральні (бокобочні), що складаються з одного тяжа флоеми, до якого щільно примикає тяж ксилеми. У стеблі флоема звернена до периферії, у листі – до нижньої сторони платівки.
Біколатеральні (двобокобочні), в яких ксилема знаходиться між двома тяжами елементів флоеми, зовнішнім і внутрішнім.
Концентричні, у яких або ксилема з усіх боків оточує флоему або флоема оточує ксилему (амфікрибральний пучок).
Складні радіальні, у яких тяжи флоеми чергуються з радіальними тяжами ксілеми, що утворюють на поперечному зрізі більш-менш зірчасту фігуру. Ці пучки характерні коріння. Пучки, що складаються з якоїсь однієї тканини - флоеми або ксілеми, називають простими або неповними. Ними закінчуються, наприклад, дрібні відгалуження жилок у листових пластинках.
Якщо флоема і ксилема розділені камбієм, в результаті діяльності якого виникають вторинні провідні тканини, пучок називають відкритим, тобто здатним до подальшій освітіклітин. Відкриті пучки властиві стеблам і корінням дводольних і голонасінних рослин, вони можуть бути колатеральними та біколатеральними. Закриті провідні пучки, що не мають камбію, характерні для стебел і коренів папоротеподібних та однодольних рослин та для більшості листя. Закриті пучки представлені концентричними, колатеральними та радіальними пучками. Пучки, що проводять, нерідко армовані склеренхімою, яка оточує пучок з усіх боків або утворює тяжи з боку флоеми або деревини. Такі пучки нерідко називають судинно-волокнистими.
Тканини, що складаються з однорідних паренхімних клітин, які заповнюють простір між іншими тканинами, називають основними. Живі клітини основної паренхіми перебувають у стані тургору й у контакті коїться з іншими тканинами збільшують механічну міцність рослини. Основна паренхіма може спеціалізуватись на виконанні різних функцій. До системи основних тканин відносяться запасні, у яких відкладається запас поживних речовин; асиміляційні, у яких відбувається фотосинтез; видільні, що містять продукти відкидання; тканини поглинання води та ґрунтових розчинів. До останніх відноситься епіблема або різодер-ма. Вона розташована на самій периферії молодого кореня та поряд зі своєю основною функцією відіграє роль покривної тканини.
У рослинах виділяють систему провітрювання, яка представлена продихами, розташованими в епідермісі, чечевичками, що знаходяться в перидермі, повітроносними порожнинами і міжклітинниками.
Вивчення будови тканин доцільно розпочати з розгляду покривних, механічних та провідних.
Різноманітність тканин у стеблі гарбуза звичайного (Cucurbita реро L.)
Шматочки стебел довжиною 2-3 см, вирізані з міжвузля з невеликою внутрішньою повітряною порожниною, фіксують спиртом наприкінці вегетаційного періоду (у серпні – вересні).
Мал. 1. Схема поперечного зрізу стебла гарбуза: еп – епідерміс, кіль – колленхіма, п – паренхіма, скл – склеренхіма, фл – флоема, кмб – камбій, кс – ксилема, ст. п. - повітряна порожнина
Стебло гарбуза в обрисі округле або округло-п'ятикутне, з п'ятипроменевою повітряною порожниною (рис. 1). Між променями порожнини розташовані п'ять великих пучків, що проводять, які добре видно неозброєним оком. Проти променів порожнини, трохи ближче до периферії стебла, знаходиться друге кільце з п'яти таких, але дрібніших пучків.
Внутрішня будовастебла вивчають на поздовжньому та поперечному зрізах. Поперечний зріз повинен захопити не менше половини перерізу стебла. Якщо такий зріз не виходить досить тонким, то для роботи з великим збільшенням мікроскопа слід приготувати ще один якомога тонший зріз, на якому повинні бути зовнішня частина стебла і хоча б один великий пучок, що проводить. Поздовжній радіальний зріз, що проходить посередині великого пучка, краще робити з міжвузля товстих стебел. Перед виготовленням зрізу стебло розрізають уздовж діаметром. Тканини, які розташовані зовні від пучка, можна видалити. Щоб зрізи не були дуже довгими, поверхню, з якої їх роблять, підсікають бритвою на відстані 0,5 см від верхнього краюшматочка. Найбільш вдалі зрізи кладуть у розчин йоду водному розчинііоди-стого калію і накривають покривним склом. Деякі зрізи можна послідовно обробляти фло-роглюцином та соляною кислотоюі розглядати їх у гліцерині.
Загальний план розташування тканин вивчають на поперечному зрізі стебла при малому збільшенні мікроскопа, будову окремих тканин розглядають при великому збільшенні на поперечному та поздовжньому зрізах.
Стебло покрите епідермісом, на поверхні якого добре видно тонку світлу плівку - кутикулу. Епідерміс складається з одного шару щільно зімкнутих живих клітин із потовщеною зовнішньою стінкою. Деякі клітини утворюють багатоклітинні волоски.
Під епідермісом розташовані ділянки механічної тканини – коленхіми. Найбільші їх перебувають у ребрах стебла. Колленхіма складається з живих багатокутних клітин із нерівномірно потовщеними стінками. Стінки потовщені в кутах, тобто в місцях з'єднання кількох клітин. Така коленхіма називається кутовою (рис. 2, А). На поперечному зрізі потовщення мають вигляд трикутників, якщо з'єднуються три клітини, або ромбів, якщо з'єднуються чотири клітини. Потовщені оболонки внаслідок сильного заломлення світла виглядають блискучими, вони добре помітні навколо округлих, гемніших, заповнених вмістом порожнин клітин. На поздовжньому зрізі клітини колінхіми подовжені.
За часом виникнення коленхіму - сама рання механічна тканина, оболонки її клітин еластичні, тому коленхіма може розтягуватися разом із зростанням стебла. Механічну роль ця тканина виконує лише в стані тургору, так як незтовщені місця оболонок при підв'язанні спадаються.
Між ділянками коленхіми, безпосередньо під нею і між іншими спеціалізованими тканинами і пучками, що проводять, розташовані тонкостінні живі, майже округлі клітини з невеликими міжклітинниками. Це паренхіма. Розміри клітин паренхіми в різних частинахстебла неоднакові.
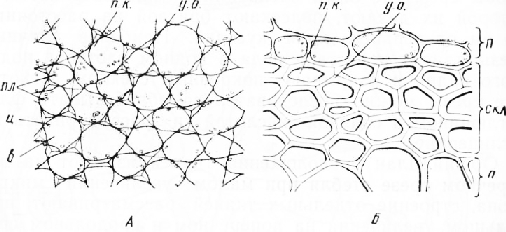
Мал. 2. Механічні тканини стебла гарбуза. А - кутова коленхіма; Б – склеренхіма: л – паренхімні клітини, у. о. - потовщення оболонки, пл - пластиди, Ч - цитоплазма, в - вакуоля, скл - склеренхіма, п. к. - порожнини клітин
У деяких паренхімних клітинах відкладається запасний крохмаль. У периферичних шарах стебла клітини основної паренхіми містять хлороплаети і виконують асиміляційну функцію.
Паренхіма, що підстилає ділянки коленхіми, межує з кількома рядами щільно зімкнутих багатокутних клітин з рівномірно потовщеними здерев'янілими стінками. Це склеренхіма (рис. 2, б). Оболонки її клітин після взаємодії з йодом набувають яскраво-оранжевого кольору, а після флороглюцинової реакції вони стають малиново-червоними. На поперечному зрізі склеренхіма розташовується кільцем. У старих стеблах клітини склеренхіми мертві, у молодших стеблах її клітинах видно залишки вмісту. Склеренхіма представлена сильно витягнутими, загостреними на кінцях волокнами, добре видно на поздовжньому зрізі.
Склеренхіма виконує механічну функцію. Вона утворюється пізніше коленхіми. Здеревніння стінок склеренхімних волокон починається після закінчення росту стебла в довжину. Оскільки здеревнілі стінки втрачають еластичність, при потовщенні стебла кільце склеренхіми нерідко розривається. Розриви заповнюються парен-хімною тканиною.
Розташування механічних тканин - коленхіми та склеренхіми - на периферії стебла підвищує опір стебла вигину, стиску та розтягуванню, яким піддаються головним чином зовнішні зони органу.
Крім найпоширенішої кутової коленхіми у рослин нерідко зустрічається коленхіма іншої будови. Так, у стеблах із сильним вторинним потовщенням розвивається коленхіма, у якої потовщені тангентальні стінки клітин, тобто стінки, паралельні поверхні органа. Радіальні стінки залишаються тонкими. Таку коленхіму називають пластинчастою (рис. 28, А). Її можна бачити в стеблі соняшника, в пагонах багатьох деревних рослин.
![]()
Мал. 3. Поперечний зріз коленхіми. А – пластинчаста; Б - пухка: п. о. первинна оболонка, в. о.-потовщена оболонка, п. к. - порожнини клітин із вмістом, пл - пластиди, мж - міжклітини
Колленхіма, в якій є міжклітини, називається пухкою (рис. 28, Б). У ній сильно потовщені стінки, що обмежують міжклітинник. Потовщені ділянки оболонок мають вигляд кілець. На зрізах, оброблених розчином йоду у водному розчині йодистого калію, міжклітини добре відрізняються від клітин клітин відсутністю вмісту. Пухка коленхіма характерна для черешків ревеню (Rheum), стебел горця (Polygonum), марі білої (Chenopodium album L.)
Будова пучків, що проводять, у стеблі гарбуза (Cucurbita реро L.)
Усі десять пучків стебла гарбуза влаштовані однаково, тому можна обмежитися розглядом одного великого пучка.
У середній частині пучка розташована ксилема або деревина. У ній навіть неозброєним оком добре помітні великі округлі порожнини клітин з здерев'янілими оболонками. Це судини, чи трахеї, - елементи, які проводять воду.
Зовнішня і внутрішня частини пучка складаються з досить дрібних елементів, що не дерев'яні, сукупність яких становить флоему, або луб. Між зовнішньою флоемою та ксилемою лежать живі тонкостінні клітини освітньої тканини – камбію. Такий пучок називається відкритим біколатераль-ним.
Ознайомившись з загальним планомрозташування провідних тканин у пучку, слід перейти до їх детального вивчення при великому (400-600 разів) збільшенні мікроскопа.
Флоема
Флоема, або луб, є складною тканиною, до складу якої поряд з провідними елементами можуть входити клітини паренхіми і флоемні волокна (у гарбуза волокон немає).
Проводять елементи флоеми - ситоподібні трубки - утворюються з подовжених, що становлять вертикальний ряд клітин, що належать прокамбію або виникли внаслідок поділу клітин камбію. Кожна клітка цього ряду ділиться поздовжньою перегородкою. Одна із сестринських клітин розростається завширшки, у її поперечних стінках з'являються дрібні наскрізні отвори - перфорації, або прориви, через які проходять тяжи цитоплазми, що з'єднують протопласти сусідніх по вертикалі клітин.
Перегородки з наскрізними отворами одержали назву ситоподібних платівок. Вертикальний ряд клітин (членників) із ситоподібними пластинками становить ситоподібну трубку. Оболонки ситоподібних трубок неодревеснілі, порівняно тонкі. Вміст представлений постінним шаром цитоплазми та слизовим клітинним соком. У функціонуючих трубках ядра зазвичай руйнуються.
Друга клітина, що відокремлювалася від материнської одночасно з клітиною-члеником ситовидної трубки, залишається вузькою: найчастіше вона ділиться кілька разів поперечними перегородками, утворюючи тяж супроводжуючих клітин, або клітин-супутниць. Протягом усього життя клітини, що супроводжують, зберігають ядро і густу цитоплазму.
Для повного уявлення про будову флоеми необхідно розглянути як поперечні, а й поздовжні зрізи. Структура зовнішньої та внутрішньої флоеми у гарбуза однакова. На поперечному зрізі флоеми ситоподібні трубки найбільш широкопросвітні, в контурі округлі або округло-багатокутні. Якщо ситоподібні трубки перерізані на рівні ситовидної пластинки, то на поперечному зрізі ця пластинка з наскрізними отворами видно в плані, а клітини, що супроводжують, здебільшого не видно. Якщо ситоподібна трубка перерізана посередині членика, то тоді дуже добре видно щільно примикають до ситоподібних трубок дрібні клітини, що супроводжують, з густим зернистим вмістом.
Ситоподібні пластинки в ситоподібних трубках можуть бути горизонтальними або трохи похилими. На строго радіальному зрізі стебла гарбуза ситоподібні платівки зазвичай розташовані майже на одному рівні, так як членики ситоподібних трубок, що знаходяться в одному радіальному ряду, є похідними однієї камбіальної клітини. У перерізаних ситоподібних платівках видно тонкі наскрізні канали. На бічних стінках ситовидних трубок зустрічаються невеликі округлі ситовидні поля, через які проходять найтонші цитоплазматичні тяжі, що з'єднують протопласти двох розташованих трубок. Порожнини ситоподібних трубок зазвичай заповнені вмістом, що згорнувся при фіксації. У ньому багато білків, тому після обробки зрізів йодним розчином вміст стає жовто-жовтогарячим. На поздовжньому зрізі вміст ситоподібних трубок має вигляд тяжів, що перетинають клітини і розширюються у ситоподібних пластинок.
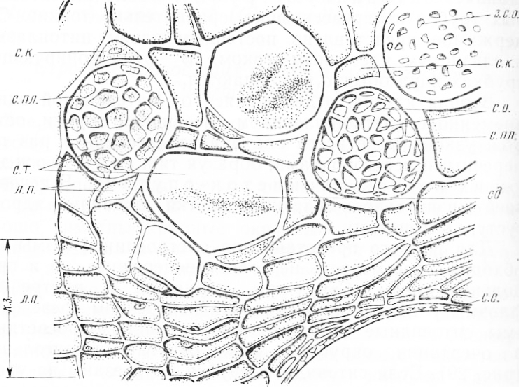
Мал. 4. Поперечний зріз флоеми та камбіальної зони стебла гарбуза: с. т. – ситоподібні трубки, с. пл. - ситоподібна платівка, с. «. - Супровідна клітина, л. л.-луб'яна паренхіма, с. о. - ситоподібні отвори, з. с. о. - закупорені калозою ситоподібні отвори, сд - вміст ситовидної трубки, к. з. - Камбіальна зона, с. с. - стінка судини
Якщо зріз пройшов косо по відношенню до поздовжньої осі стебла, то ситовидні пластинки мають вигляд еліпса з овальними наскрізними отворами, а перерізані тяжи вмісту, що згорнувся, помітні як темні згустки лише у ситовидних пластинок.
У міру старіння ситоподібних трубок ситоподібні прориви поступово закупорюються особливою речовиною - калозою, при цьому отвори звужуються і на поперечному зрізі фпоеми мають вигляд невеликих точок. До кінця вегетаційного періоду каллоза утворює мозолисті тіла, які являють собою блискучі потовщення, що сильно заломлюють світло, на обох сторонах ситовидної пластинки.
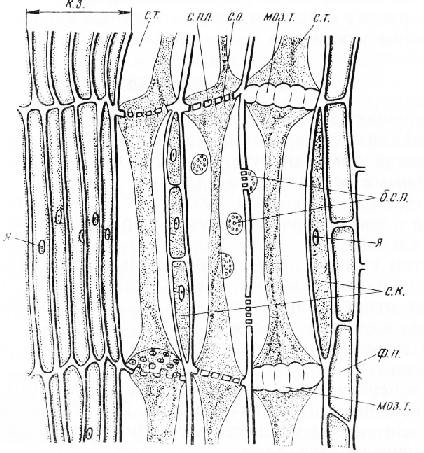
Мал. 5. Поздовжній зріз флоеми та камбіальної зони стебла гарбуза: с. т. – ситоподібні трубки з тяжами вмісту, с. пл. - ситоподібні платівки, с. о.- ситоподібні отвори, б. с. п.- ситоподібні поля на бічних стінках, с. к. - супровідні клітини, ф. п. – флоемна паренхіма, моз. т. - мозолисте тіло, к. з. - камбіальна зона, я - ядро
Мозолисті тіла особливо добре помітні на поздовжніх зрізах старої флоеми, найбільш віддаленої від камбію.
Уздовж членика ситовидної трубки, щільно прилягаючи до нього, розташована одна довга або тяж з кількох коротких клітин, що супроводжують.
Загострені кінці крайових клітин тяжа знаходяться на рівні ситоподібних пластинок, що вказує на спільність походження клітин, що супроводжують, і члеників ситоподібних трубок.
Живі паренхімні клітини, що входять до складу флоеми, відрізняються від члеників ситоподібних трубок меншими розмірами і відсутністю сит, але вони значно більші за супроводжуючі клітини, сильніше вакуолізовані і здаються світлішими.
Камбіальна зона
У пучку, що проводить, між зовнішньою флоемою і ксилемою знаходяться живі тонкостінні клітини, що становлять камбіальну зону. Клітини розташовані правильними радіальними рядами.
Усі клітини цієї зони виникають у результаті поділу клітин однорядного шару освітньої тканини – камбію.
Клітини камбію діляться поздовжніми тангентальними (тобто паралельними поверхні органу) перегородками, відкладаючи клітини назовні, у бік флоеми, і всередину, у бік ксилеми.
Чим інтенсивніший поділ камбію, тим ширша камбіальна зона.
На поперечному зрізі клітини цієї зони мають майже прямокутні контури. Тангентальні стінки клітин, що у сусідніх радіальних рядах, зазвичай не збігаються.
На поздовжньому зрізі (рис. 30) клітини розташовані ярусами. Вони витягнуті в довжину і мають слабко загострені або трохи закруглені закінчення, що знаходяться на одному рівні. Поздовжні стінки клітин одного ярусу вклинюються між стінками клітин сусіднього вертикалі ярусу. У клітині видно цитоплазму та ядро.
Периферичні клітини камбіальної зони згодом диференціюються елементи вторинної флоеми. На успішно зроблених поздовжніх радіальних зрізах можна побачити, що довжина клітин камбіальної зони майже дорівнює довжині члеників ситоподібних трубок. З внутрішніх клітинкамбіальної зони формуються елементи вторинної ксилеми.
Ксилема
Ксилема (деревина) складається з судин (трахей) з досить товстою здеревілою оболонкою. Найбільш широкопросвітні судини видно навіть неозброєним оком. Між судинами розташовані дрібні клітини, в контурі багатокутні.
Завдання.
1. При малому збільшенні мікроскопа замалювати схему будови поперечного зрізу стебла гарбуза, відзначивши на ній епідерміс, ділянки коленхіми, паренхіму, кільце склеренхіми, біколатеральні пучки з їх тканинами і центральну повітряну порожнину. При замальовці схеми необхідно дотримуватись масштабу. Найкраще починати схему з обрисів повітряної порожнини і контурів пучків, що проводять. Це можна зробити, розглядаючи зріз під лупою. Обриси інших тканин відзначають, розглядаючи зріз при мінімальному збільшенні мікроскопа, пересуваючи препарат. Щоб схема була наочною, для кожної тканини вибирають. умовні позначення(точки, одностороннє або двостороннє штрихування тощо). Можна скористатися і кольоровими олівцями. Основну тканину не зафарбовують, флоему можна позначити синім, ксилему – червоним кольором, кружальцями у ній показують найбільші судини. Схема має бути забезпечена пояснювальними написами. Лінії, які виносять написи, бажано робити горизонтальними; якщо це чомусь неможливо, потрібно намагатися не перехрещувати лінії.
2. При великому збільшенні мікроскопа з поперечного зрізу детально замалювати клітини епідермісу, коленхіми, склеренхіми, флоеми та камбіальної зони. У флоемі зобразити перерізану ситоподібну трубку з внутрішнім вмістом, ситоподібну пластинку в плані, що супроводжують і паренхімні клітини.
3. З поздовжнього зрізу замалювати при великому збільшенні мікроскопа: а) ситоподібну трубку з супроводжуючими клітинами, відзначивши в ній перерізану ситоподібну пластинку, тяжі згорнутого вмісту, мозолисте тіло, ситоподібні поля на бічних стінках; б) клітини камбіальної зони.
Будова елементів ксилеми, що проводять, в стеблі соняшника однорічного (Helianthus annuus L.)
Ксилема, або деревина, так само як і флоема, є складною тканиною, що складається не тільки з елементів, що проводять воду з розчиненими в ній. мінеральними речовинами, але і з клітин, що виконують механічну та запасну функції.
До провідних елементів ксилеми відносяться судини, або трахеї, і трахеїди. Трахеїди – це довгі прозенхімні клітини з сильно скошеними кінцями та численними облямованими порами на бічних стінках. Судини - порожнисті трубки, що є вертикальним рядом подовжених клітин (членників). Сумежні стінки, що відокремлюють одну клітину вертикального ряду від іншої, у процесі розвитку судин руйнуються, на їх місці виникають наскрізні отвори – перфорації. Бічні стінки члеників товщають і здерев'янюють, протопласти клітин відмирають.
Особливості будови провідних елементів найкраще видно на поздовжніх зрізах.
Шматочки стебла соняшнику товщиною близько 1 см, вирізані з міжвузля і фіксовані спиртом, розрізають уздовж по радіусах і очищають від паренхімних клітин серцевини. Потім роблять тонкі радіальні зрізи твердої частини стебла або серію тангентальних зрізів, внутрішньої зони деревини, що межує із серцевиною. Зрізи спочатку обробляють спиртовим розчином флороглюцину, а потім міцною соляною кислотою і укладають у гліцерин. Їх розглядають за великого збільшення мікроскопа.
Судини, розташовані поблизу серцевини, мають тонку первинну целюлозну оболонку і здерев'янілу вторинну оболонку у вигляді окремих кілець. Після проведення реакції на здеревнення кільця добре помітні завдяки червоному забарвленню. Ці судини називають кільчастими.
Поруч із ними, ближче до периферії стебла, знаходяться судини з вторинним потовщенням оболонки у вигляді одинарної або подвійний спіралі. Це спіральні судини. Завитки спіралей окремих клітин-членників, що складають судину, з'єднуючись кінцями, утворюють єдину
суцільну спіраль. На препаратах спіральні та кільчасті судини можуть бути розрізані вздовж, тоді їх потовщення мають вигляд напівкілець. Іноді в одній посудині спіральні потовщення чергуються з кільчастими. Цей проміжний типсудин називають кільчасто-спіральним.
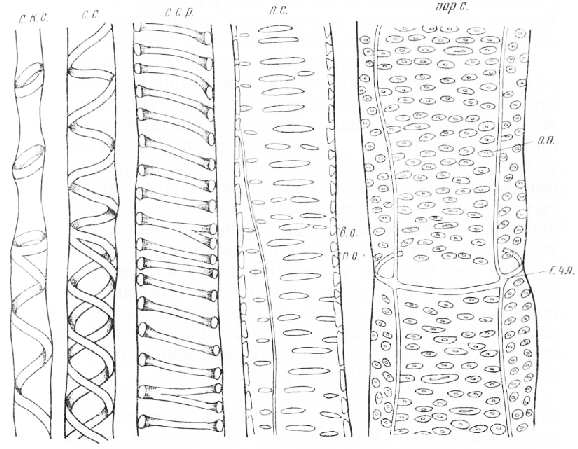
Мал. 6. Посудини на поздовжньому зрізі стебла соняшнику: с. к. с. - спірально-кільчаста судина, с. с. - спіральна судина, с. с. нар. - спіральний посуд у розрізі, л. с. - Сходовий посуд, пор. с. - пористий посуд, п. о. - первинна оболонка, ст. о. - Вторинна оболонка, о. п. - облямовані пори, р. чл. - кордон членика судини
Кільчасті, спіральні та спірально-кільчасті судини виникають в онтогенезі стебла дуже рано, коли зростання органу в довжину ще не закінчено. Вони формуються з клітин прокамбію і належать протоксилемі. У міру зростання стебла непотовщена первинна оболонка цих судин розтягується, а кільчасті потовщення та завитки спіральних потовщень розсуваються. Згодом під тиском сусідніх клітин незтовщені стінки кільчастих судин здавлюються, їх діаметр у цих місцях зменшується.
До периферії від спіральних судин розташовані судини з більш потовщеною оболонкою, не здатною до розтягування. Ті з них, що розвиваються з прокамбію, відносяться до метаксилеми. Протоксилема та метаксилема становлять первинну ксилему. Посудини, що утворюються пізніше, внаслідок діяльності камбію, входять до складу вторинної ксилеми. Судини метаксилеми та вторинної ксилеми мають сходові та сітчасті потовщення бічних стінок.
Формуванню сходової судини передує утворення спіральних потовщень вторинної оболонки, причому окремі обороти спіралей настільки зближені, що в деяких місцях вони з'єднуються. Вузькі простори між нез'єднаними ділянками спіральних потовщень є пори. У плані вони мають вигляд витягнутих по ширині судини довгастих або майже щілинних ділянок, розташованих вертикальними рядами, як сходинки. Такий тип порівності стінок судини називається сходовим.
При сітчастому потовщенні пори в стінках судини округлі або овальні в контурі та розташовані в один або кілька рядів. У стеблі соняшнику ці судини знаходяться зовні від сходових. Численні, дуже дрібні, які щільно примикають одна до одної пори можна було бачити в судинах гарбуза.
Будова пір судин відрізняється від будови простих пір в оболонках паренхімних клітин наявністю облямівки. Облямуванням називається піднесеність вторинних шарів оболонки над поровим каналом (докладніше про будову облямованих пір див. розділ «Деревина хвойних рослин»). Пори на поздовжніх стінках судин служать щодо води у горизонтальному напрямі. Якщо посудина в поперечному перерізібагатокутний, то на поздовжньому зрізі на його стінці помітні вертикальні смуги.
Велика довжинаяк наслідок руйнування поперечних стінок між клітинами, потовщені одревеснілі бічні стінки, Що забезпечують міцність, відсутність живого вмісту характеризує судини як найбільш високоорганізований тип провідних елементів, найкращим чиномвиконують функцію проведення води.
Крім судин до складу ксилеми входять живі тонкостінні паренхімні клітини, що розташовуються між судинами, а також волокна, що сильно здеревніли, подібні за будовою з волокнами склеренхіми.
Завдання. При великому збільшенні мікроскопа замалювати кільчасті, спіральні, сходові та пористі судини. Звернути увагу на характер потовщень стінок, контур і розташування пір, відзначити перфорацію.


