Вакуоль функції. Вакуолі та клітинний сік
Вакуолі та клітинний сік
Більшість зрілих клітин рослин характерна велика центральна вакуоля, що займає до 70-90% обсягу клітини. При цьому протопласт з усіма органелами розташовується у вигляді дуже тонкого пісенного шару, що вистилає клітинну стінку. У постінному протопласті зазвичай трапляються дрібні цитоплазматичні вакуолі. Іноді ядро розташовується в центрі клітини ядерній кишеньціцитоплазми, який пов'язаний з постінним шаром найтоншими цитоплазматичними тяжами, що перетинають центральну вакуоль.
Клітинний сік є водний розчин різних речовин, що є продуктами життєдіяльності протопласту, в основному, запасними речовинами та покидьками. Реакція клітинного соку зазвичай слабокисла чи нейтральна, рідше лужна. Речовини, що входять до складу клітинного соку, надзвичайно різноманітні. Це вуглеводи, білки, органічні кислоти та їх солі, амінокислоти, мінеральні іони, алкалоїди, глікозиди, таніди, пігменти та інші розчинні у воді сполуки. Більшість із них належить до групи ергастичних речовин – продуктів метаболізму протопласту, які можуть з'являтися та зникати у різні періоди життя клітини. Багато речовин клітинного соку утворюються лише у рослинних клітинах.
Вуглеводи клітинного соку рослин представлені моносахаридами (глюкоза, фруктоза), дисахаридами (сахароза) та полісахаридами (слизу, інулін).
Глюкоза (виноградний цукор) та фруктоза (плодовий цукор) накопичуються в великих кількостяху соковитих плодах. Сахароза (буряковий цукор) у великих кількостях накопичується в коренеплодах цукрових буряків та стеблах цукрової тростини. Для ряду сімейств рослин (кактусові, товстянкові, орхідні) характерне накопичення в клітинному соку слизів, що утримують воду. Інулін – запасний полісахарид, що відкладається у вигляді колоїдного розчинуу клітинному соку підземних органів складноцвітих замість крохмалю.
Білки накопичуються у вигляді колоїдного розчину у вакуолях клітин насіння, що дозріває. При зневодненні насіння на пізніх етапах розвитку вода видаляється з вакуолей, концентрація білка в клітинному соку підвищується, і він перетворюється на стан твердого гелю. Дегідратовані вакуолі зрілого насіння називають алейроновими зернами.
З органічних кислоту клітинному соку найчастіше зустрічаються лимонна, яблучна, бурштинова та щавлева. Ці кислоти знаходяться у великій кількості у клітинному соку незрілих плодів, надаючи їм кислого смаку. При дозріванні плодів органічні кислоти можна використовувати як субстрати дихання, тому кислий смак плодів зазвичай зникає. Солі органічних кислот разом із мінеральними іонами відіграють велику роль в осмотичних процесах.
Таніди (дубільні речовини) – полімерні фенольні сполуки терпкого смаку. Вони мають антисептичні властивості і захищають тканини рослин від інфекцій та загнивання. Особливо багаті на дубильні речовини клітини кори стебел і коренів (дуб, верба), незрілих плодів (волоський горіх), листя (чай) та деяких патологічних наростів – галлів. Таніди використовуються в медицині, для дублення шкіри, забарвлення тканини у темно-коричневий колір.
Алкалоїди – різноманітні в хімічному відношенніазотовмісні органічні речовини, що мають гіркий смак. Вони мають властивості підстав і містяться в клітинному соку, як правило, у вигляді солей. Багато алкалоїдоносних рослин отруйні і не поїдаються травоїдними тваринами. У клітинах, що містять алкалоїди, не розвиваються суперечки та зачатки мікроорганізмів, рослини не уражаються грибними та бактеріальними хворобами. Особливо багаті на алкалоїди представники сімейств пасльонових, макових, маренових, лютикових та ін.
Глікозиди – велика групаприродних речовин, сполуки цукрів зі спиртами, альдегідами, фенолами та іншими речовинами Ряд глікозидів рослин використовують у медицині. До глікозидів належать також пігменти клітинного соку. флавоноїди. Один з них - антоціани- Надають клітинному соку червоний, синій або Фіолетовий колір; інші – флавони– жовтий. З антоціанами пов'язане фарбування квіток багатьох рослин. Колірна гама обумовлена реакцією клітинного соку: якщо вона кисла, то панують червоні тони, нейтральна – фіолетові, при слаболужній реакції – сині. На виникнення відтінків впливає також утворення антоціанами комплексів з різними металами. Флавони зумовлюють жовтий колірпелюсток ряду рослин.
Значення органічних кислот, танідів, алкалоїдів та глікозидів клітинного соку в обміні речовин клітини з'ясовано недостатньо. Раніше їх розглядали як кінцеві продукти обміну. В даний час показано, що багато з них можуть знову залучатися до процесів метаболізму і тому їх можна розглядати і як запасні речовини.
Крім функції накопичення запасних речовин та покидьків, вакуолі в рослинних клітинах виконують ще одну важливу функцію- Підтримка тургора. Концентрація іонів та цукрів у клітинному соку центральної вакуолі, як правило, вище, ніж у клітинній стінці; Тонопласт значно уповільнює дифузію з вакуолі цих речовин і водночас легко проникаємо для води. Тому вода надходитиме у вакуолю. Такий односпрямований процес дифузії води через вибірково проникну мембрану зветься осмосу. Вода, що надходить у клітинний сік, надає тиск на постінний протопласт, а через нього і на клітинну стінку, викликаючи напружений, пружний її стан, або тургорклітини. Тургор забезпечує збереження неодревесневшими органами рослини форми та положення у просторі, а також їх опір дії механічних факторів.
Якщо клітину помістити в гіпертонічний розчин якоїсь нетоксичної солі або цукру (тобто в розчин більшої концентрації, ніж концентрація клітинного соку), відбувається осмотичний вихід води з вакуолі. Внаслідок цього її обсяг скорочується, еластичний постінний протопласт відходить від клітинної стінки, тургор зникає, настає плазмолізклітини ( рис.2.9).
Рис. 2.9. Схема плазмолізу: 1 – клітина у стані тургору; 2 – початок плазмолізу; 3 – повний плазмоліз.
Плазмоліз зазвичай оборотний. При поміщенні клітини у воду або гіпотонічний розчин вода знову енергійно поглинається центральною вакуоллю, протопласт знову притискається до клітинної стінки, тургор відновлюється. Плазмоліз може бути показником живого стану клітини, мертва клітина не плазмолізується, тому що не має вибірково проникних мембран.
Втрата тургору викликає зав'ядання рослини. При зав'яданні повітря в умовах недостатнього водопостачання тонкі стінки клітин зморщуються одночасно з протопластом і робляться складчастими.
Тургорний тиск не тільки підтримує форму неодревесневших частин рослин, він є також одним із факторів росту клітини, забезпечуючи зрістклітин розтягуванням, тобто за рахунок поглинання води та збільшення розміру вакуолі. У тварин клітин центральна вакуоля відсутня, їх зростання відбувається головним чином за рахунок збільшення кількості цитоплазми, тому розмір тварин клітин зазвичай менше, ніж рослинних.
Центральна вакуоля виникає шляхом злиття численних дрібних вакуолей, які є у меристематичних (ембріональних) клітинах. Ці цитоплазматичні вакуолі утворюються, як вважають, за рахунок мембран ендоплазматичної мережічи апарату Гольджі.
Увімкнення
Утворення включень викликане надлишковим накопиченням деяких продуктів обміну речовин у певних ділянкахклітини – у вакуолі, гіалоплазмі, різних органелах, рідше у клітинній стінці. Ці речовини часто випадають в осад в аморфному вигляді або у формі кристалів. включень. Включення мають певну формуі добре видно світловий мікроскоп. За наявності тих чи інших включень, їх формі та розподілу можна відрізнити одні види, пологи та сімейства рослин від інших, тому вони часто є важливою діагностичною ознакою при аналізі лікарської рослинної сировини.
Включення є або запасні речовини(тимчасово виведені з обміну речовин сполуки), або кінцеві продуктиобміну. До першої категорії включень відносяться крохмальні зерна,ліпідні крапліі відкладення білків; до другої – кристалидеяких речовин.
Крохмальні зерна- Найбільш поширені включення рослинних клітин. Полісахарид крохмаль- Основний тип запасних поживних речовинрослин. Він є і самим важливим з'єднанням, що використовуються в їжу рослиноїдними тваринами. Крохмаль зерновок хлібних злаків, бульб картоплі, плодів банана – найважливіше джерелохарчування людей. Пшеничне борошно складається із зерен крохмалю майже на 75%, у бульбах картоплі крохмаль становить 20-30%. У хімічному відношенні крохмаль є альфа-1,4-D-глюкан, молекули мають вигляд розгалужених ланцюгів, в крохмальному зерні вони розташовуються по радіусах.
Крохмальні зерна утворюються в стромі пластид. У хлоропластах на світлі відкладаються зерна асиміляційного (первинного) крохмалю, що утворюються при надлишку цукрів – продуктів фотосинтезу Освіта осмотично неактивного крохмалю запобігає підвищенню осмотичного тиску в хлоропласті. Вночі, коли фотосинтез не відбувається, асиміляційний крохмаль за допомогою ферментів гідролізується до цукрів та транспортується до інших частин рослини. Запасний (вторинний) крохмальвідкладається в амілопластах клітин різних органіврослин (коріння, підземні пагони, насіння) з цукрів, що притікають з фотосинтезуючих клітин. При необхідності запасний крохмаль також перетворюється на цукру.
Утворення крохмальних зерен починається в певних точках строми пластиди, які називаються освітніми центрами. Зростання зерна відбувається шляхом послідовного відкладення шарів крохмалю навколо освітнього центру. Сумежні шари в одному зерні можуть мати різний показник заломлення світла, і тоді вони помітні під мікроскопом. шаруватікрохмальні зерна. Розташування шарів може бути концентричним(пшениця) або ексцентричним(картопля) ( Рис. 2.10). Якщо в амілопласті є один освітній центр, навколо якого відкладаються шари крохмалю, виникає простоїзерноякщо два і більше – то утворюється складнезерно, Що складається ніби з кількох простих. Напівскладне зерноутворюється в тому випадку, якщо крохмаль спочатку відкладається навколо декількох точок, а потім, після дотику простих зерен, навколо них виникають загальні шари ( Рис. 2.10).
Форма, розмір, кількість в амілопласті та будова (становище освітнього центру, шаруватість, наявність або відсутність тріщин) крохмальних зерен часто специфічні для виду рослини ( Рис. 2.10). Зазвичай крохмальні зерна мають сферичну, яйцеподібну або лінзовидну форму, проте у картоплі вона неправильна. Найбільші зерна (до 100 мкм) характерні для клітин бульб картоплі, у зернівці пшениці вони двох розмірів – дрібні (2-9 мкм) і більші (30-45 мкм). Для клітин зернівки кукурудзи характерні дрібні зерна (5-30 мкм). Складні крохмальні зерна у рису, вівса, гречки.
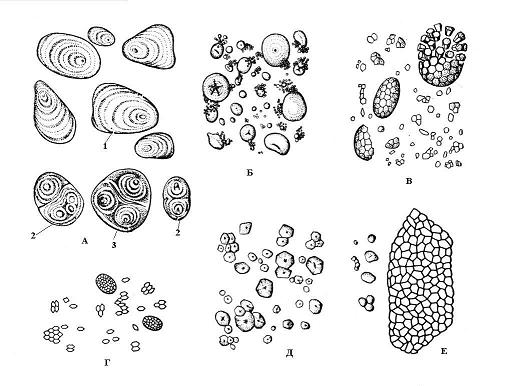
Рис. 2.10. Крохмальні зерна різних видіврослин: А - картопля; Б – пшениця; В – овес; Г – рис; Д – кукурудза; Е – гречка; 1 – просте зерно; 2 – складне зерно; 3 – напівскладне зерно.
Реактивом на крохмаль є розчин йоду в розчині йодиду калію – реактивЛюголя. Він фарбує крохмальні зерна у синьо-фіолетовий колір.
Відкладення крохмалю широко поширені у всіх органах рослини, але особливо багате на нього насіння, підземні пагони (полуниці, цибулини, кореневища), паренхіма провідних тканин коренів і стебел деревних рослин.
Ліпідні краплізустрічаються практично у всіх рослинних клітинах. Жирні олії накопичуються у величезної кількостірослин і за своїм значенням є другою після крохмалю формою запасних поживних речовин. Особливо багаті на них насіння та плоди. Насіння деяких рослин (соняшник, бавовник, арахіс) може містити до 40% олії від маси сухої речовини. Тому рослинні жири одержують, головним чином, із насіння.
Ліпідні краплі накопичуються безпосередньо у гіалоплазмі. Вони виглядають як дрібні сферичні тіла, кожна крапля відокремлена від гіалоплазми мембраною. Іноді ліпідні краплі називають сферосомами.
Реактивом на жирне масло є барвник суданIII, ліпідні краплі забарвлюються їм в оранжево-червоний колір.
Білкові включенняу вигляді різноманітних аморфних чи кристалічних відкладень утворюються у різних органелах клітини. Найбільш часто білкові кристали можна зустріти в ядрі, рідше - в гіалоплазмі, стромі пластид, у розширеннях цистерн ендоплазматичної мережі, матриксі пероксисом та мітохондрій. Розмір білкових кристалів найчастіше знаходиться за межами роздільної здатності світлового мікроскопа.
Запасні білки належать до категорії простих білків – протеїнів, на відміну від складних білків –протеїдів, що становлять основу протопласту У найбільшій кількостівони відкладаються в запасній тканині сухого насіння у вигляді алейронових зерен, або білковихтелець.
Алейронові зерна зазвичай мають сферичну форму та різний розмір (0,2-20 мкм). Вони оточені мембраною і містять аморфний білковий матрикс, в який занурені кристалічні включення – один (рідше, 2-3) білковий кристал ромбоедричної форми та округлі глобоїди(від одного до багатьох) ( Рис. 2.11). Глобоїди складаються з фітину (солі інозитгексафосфорної кислоти) і є місцем зберігання запасного фосфору. Алейронові зерна, що містять кристали, називають складними. Вони характерні для запасних клітин насіння олійних рослин (льон, соняшник, гарбуз, гірчиця, рицина та ін). Рідше зустрічаються простіалейронові зерна, що не містять кристалів, а лише аморфний білок (бобові, рис, кукурудза) ( Рис. 2.12).
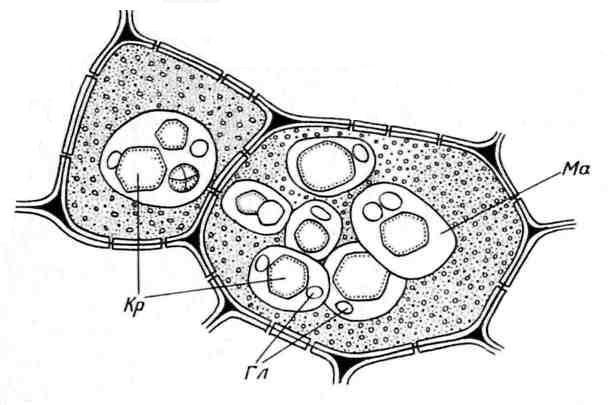
Рис. 2.11. Алейронові зерна в клітинах ендосперму насіння рицини:Кр– білкові кристали; Гол- Глобоїди; Ма– білковий матрикс.
Запасні білки під час розвитку насіння відкладаються у вакуолі. При дозріванні насіння, що супроводжується зневодненням, білкові вакуолі висихають, білок і фітин випадають з розчину в осад і можуть кристалізуватися. При проростанні насіння алейронові зерна поглинають воду, набухають і поступово перетворюються на типові вакуолі. Білки та речовини глобоїдів витрачаються на зростання та розвиток проростка.
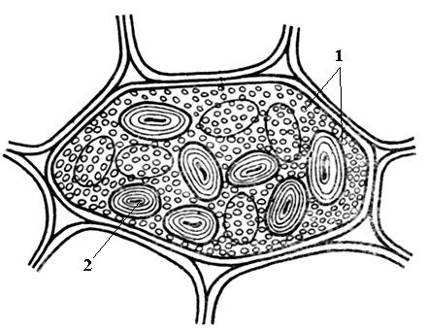
Рис. 2.12. Прості алейронові та крохмальні зерна в клітці сім'ядолі насіння квасолі: 1 - прості алейронові зерна; 2 – крохмальне зерно.
Білкові включення можна пофарбувати реактивом Люголя у золотисто-жовтий колір.
Кристалі кальцію оксалатучасто зустрічаються у рослинних клітинах. Вони відкладаються лише у вакуолях. Форма кристалів кальцію оксалату досить різноманітна ( Рис. 2.13)та часто специфічна для певних рослин, що використовується при діагностиці лікарської рослинної сировини. Це можуть бути одиночнікристалиромбоедричної, октаедричної або подовженої форми (листя блекоти), друзі– зірчасті зростки кристалів кулястої форми (листя споришу, дурману, сенни, коріння ревеню), рафіди- дрібні голчасті кристали, зібрані в пучки (листя конвалії, кореневища марени), стилоїди- Більші, паличкоподібні кристали (листя конвалії) і кристалічнийпісок- Скупчення безлічі дрібних одиночних кристалів (листя беладони). Найчастіше зустрічаються друзі. 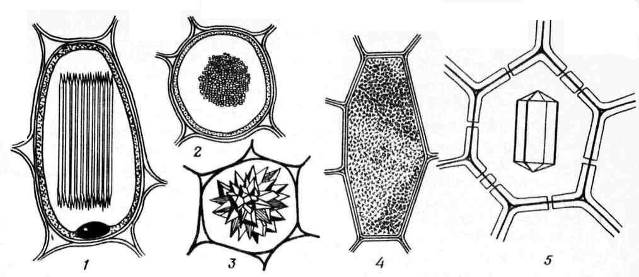
Рис. 2.13. Форми кристалів кальцію оксалату: 1,2 - рафіди (1 - вид збоку, 2 - вид на поперечному зрізі); 3 – друза; 4 – кристалічний пісок; 5 – одиночний кристал.
Уздовж волокон у корі або вздовж жилок листя у ряду рослин (кора дуба, коріння солодки, листя сени) зустрічається кристалоносна обкладка– розташовані паралельними рядамиклітини з одиночними кристалами оксалату кальцію ( рис.2.14).
![]()
Рис. 2.14. Жилка із кристалоносною обкладкою в листі сенни.
На відміну від тварин, які виділяють надлишок іонів у зовнішнє середовищеразом із сечею, рослини, що не мають розвинених органів виділення, змушені накопичувати їх у тканинах. Зазвичай вважають, що кристали кальцію оксалату кінцевий продуктжиттєдіяльності клітини, призначений для виведення надлишків кальцію. Дійсно, кристали утворюються у великих кількостях у тих органах і тканинах, які рослини іноді скидають (листя і кора). Однак, є дані, що кристали можуть зникати з вакуолей. У разі їх можна як місце відкладення запасного кальцію.
Підтвердити хімічну природу кристалів оксалату кальцію можна дією концентрованих мінеральних кислот. Під дією кислоти хлористоводневої кристали розчиняються. При дії кислоти сірчаної кальцію оксалат перетворюється на нерозчинний кальцію сульфат (гіпс), що утворює численні голчасті кристали.
До кристалічних включень близькі цистоліти. Вони найчастіше складаються з кальцію карбонату або кремнезему і являють собою гроноподібні утворення, що виникають на виступах клітинної стінки, що вдається всередину клітини ( Рис. 2.15). Цистоліти характерні для рослин сімейств кропив'яних, тутових. Значення цистолітів поки що не з'ясовано.
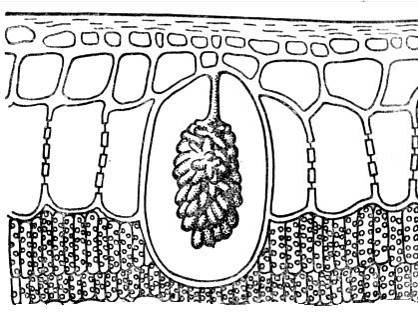
Рис. 2.15. Цистоліт у клітині епідерми аркуша фікуса.
Клітинна стінка
Клітинна стінка ( клітинна оболонка) – характерна ознакарослинної клітини, що відрізняє її від клітини тварини. Клітинна стінка надає клітині певної форми. Культивовані на спеціальних живильних середовищах клітини рослин, у яких ферментативним шляхом видаляється стінка, завжди набувають сферичної форми. Клітинна стінка надає клітині міцності та захищає протопласт, вона врівноважує тургорний тиск і перешкоджає, таким чином, розриву плазмалеми. Сукупність клітинних стін утворює внутрішній скелет, що підтримує тіло рослини і надає йому механічної міцності.
Клітинна стінка безбарвна та прозора, легко пропускає сонячне світло. Зазвичай стіни просочені водою. За системою клітинних стінок здійснюється транспорт води та розчинених у ній низькомолекулярних сполук (транспорт по апопласту).
Клітинна стінка складається в основному з полісахаридів, які можна поділити на скелетні речовиниі речовини матриксу.
Скелетною речовиноюклітинної стінки рослин є целюлоза (клітковина), Що являє собою бета-1,4-D-глюкан Це найпоширеніше органічна речовинабіосфери. Молекули целюлози є дуже довгими нерозгалуженими ланцюгами, вони розташовуються паралельно один одному групами по кілька десятків і скріплені численними водневими зв'язками. В результаті утворюються мікрофібрили, які створюють структурний каркас стінки та зумовлюють її міцність. Мікрофібрили целюлози видно лише в електронному мікроскопі, їх діаметр дорівнює 10-30 нм, довжина досягає декількох мкм.
Целюлоза нерозчинна та не набухає у воді. Вона дуже інертна у хімічному відношенні, не розчиняється в органічних розчинниках, концентрованих лугах та розведених кислотах. Мікрофібрили целюлози еластичні та дуже міцні на розрив (подібні зі сталлю). Ці властивості визначають широке застосуванняцелюлози та її продуктів. Світова продукція бавовняного волокна, що складається майже цілком із целюлози, становить 1,5 10 7 тонн на рік. З целюлози отримують бездимний порох, ацетатний шовк та віскозу, целофан, папір. Якісну реакціюна целюлозу проводять з реактивом хлор-цинк-йод, целюлозна клітинна стінказабарвлюється у синьо-фіолетовий колір.
У грибів скелетною речовиною клітинної стінки є хітін- Полісахарид, побудований з залишків глюкозаміну. Хітін ще міцніший, ніж целюлоза.
Мікрофібрили занурені в аморфний матриксзазвичай являє собою насичений водою пластичний гель. Матрикс є складною сумішшю полісахаридів, молекули яких складаються з залишків декількох різних цукрів і є більш короткими, ніж у целюлози, і розгалужені ланцюги. Матричні полісахариди визначають такі властивості клітинної стінки, як сильна набухання, висока проникність для води та розчинених у ній низькомолекулярних сполук, катіонообмінні властивості. Полісахариди матриксу ділять на дві групи. пектинові речовиниі геміцелюлози.
Пектинові речовинисильно набухають чи розчиняються у воді. Вони легко руйнуються під дією лугів та кислот. Найпростішими представниками пектинових речовин є розчинні у воді пектові кислоти– продукти полімеризації альфа-D-галактуронової кислоти (до 100 одиниць), пов'язаних 1,4-зв'язками в лінійних ланцюгах (альфа-1,4-D-галактуронан). Пектинові кислоти (пектин)– це високомолекулярні (100-200 одиниць) полімерні сполуки альфа-D-галактуронової кислоти, в яких карбоксильні групи частково метильовані. Пектатиі пектинати– кальцієві та магнієві солі пектових та пектинових кислот. Пектинові кислоти, пектати та пектинати розчиняються у воді в присутності цукрів та органічних кислот з утворенням щільних гелів.
У клітинних стінках рослин переважно присутні протопектини– високомолекулярні полімери метоксильованої полігалактуронової кислоти з арабінанами та галактанами, у дводольних рослин до складу ланцюгів галактуронану входить невелика кількість рамнози. Протопектини нерозчинні у воді.
Геміцелюлозиє розгалужені ланцюги, побудовані з залишків нейтральних цукрів, частіше зустрічаються глюкоза, галактоза, манноза, ксилоза; ступінь полімеризації 50-300. Геміцелюлози хімічно стійкіші, ніж пектинові речовини, вони важче гідролізуються і слабше набухають у воді. Геміцелюлози можуть відкладатися в стінках клітин насіння як запасні речовини (фінікова пальма, хурма). Пектинові речовини та геміцелюлози пов'язані взаємними переходами. Крім полісахаридів, в матриксі клітинних стін присутній спеціальний структурний білок. Він пов'язаний із залишками цукру арабінози і тому є глікопротеїдом.
Матричні полісахариди не просто заповнюють проміжки між целюлозними мікрофібрилами. Їхні ланцюги розташовуються впорядковано і утворюють численні зв'язки як один з одним, так і з мікрофібрил, що значно підвищує міцність клітинної стінки.
Клітинні стінки рослин часто піддаються хімічним змінам. Одервеснення, або лігніфікаціявідбувається у тому випадку, якщо в матриксі відкладається лігнін– полімерна сполука фенольної природи, нерозчинна у воді. Клітинна стінка, що здеревніла, втрачає еластичність, різко підвищується її твердість і міцність на стиск, знижується проникність для води. Реактивами на лігнін є: 1) флороглюцині концентрована хлористоводневаабо сірчана кислота(одревеснілі стінки набувають вишнево-червоного забарвлення) і 2) сульфатаніліну, під впливом якого здерев'янілі стіни стають лимонно-жовтими. Лігніфікація характерна для стінок клітин провідної тканини ксилеми (деревини) та механічної тканини склеренхіми.
Опробковування, або суберинізаціявідбувається в результаті відкладення з внутрішньої сторониклітинної стінки гідрофобних полімерів – Суберінаі воску. Суберин являє собою суміш ефірів полімерних жирних кислот. Мономерами воску є жирні спирти та воскові ефіри. Віск легко витягується органічними розчинниками та швидко плавиться, утворює кристали. Суберин – аморфна сполука, яка не плавиться і не розчиняється в органічних розчинниках. Суберин і віск, утворюючи паралельні шари, що чергуються, вистилають всю порожнину клітини з внутрішньої сторони у вигляді плівки. Суберинова плівка практично непроникна для води та для газів, тому після її утворення клітина зазвичай відмирає. Опробковування притаманно стінок клітин покривної тканини пробки. Реактивом на випробувану клітинну стінку є судан III, забарвлення оранжево-червоне.
Кутинізаціїпіддаються зовнішні стінки клітин покривної тканини епідерми. Кутіні вісквідкладаються шарами, що чергуються на зовнішній поверхні клітинної стінки у вигляді плівки – кутикули. Кутин являє собою жироподібну полімерну сполуку, близьку за хімічною природою та властивостями суберину. Кутикула оберігає рослину від зайвого випаровування води із поверхні рослини. Забарвити її можна реактивом судан IIIв оранжево-червоний колір.
Мінералізаціяклітинної стінки відбувається внаслідок відкладення в матриксі великої кількостімінеральних речовин, найчастіше кремнезему (оксиду кремнію), рідше оксалату та карбонату кальцію. Мінеральні речовининадають стінці твердість та крихкість. Відкладення кремнезему характерне для клітин епідерми хвощів, осок та злаків. Набута в результаті відокремлення жорсткість стебел і листя служить захисним засобомпроти равликів, а також значно знижує поїдання та кормову цінність рослин.
У деяких спеціалізованих клітин спостерігається ослизненняклітинної стінки. При цьому замість вторинної целюлозної стінки відбувається відкладення аморфних, сильно гідратованих кислих полісахаридів у вигляді слизіві камедей, близьких за хімічною природою до пектинових речовин Слиз добре розчиняються у воді з утворенням слизових розчинів. Камеді клейкі, витягуються у нитки. У сухому вигляді вони мають рогову консистенцію. При відкладенні слизу протопласт поступово відтісняється до центру клітини, його обсяг та обсяг вакуолі поступово зменшуються. Зрештою, порожнина клітини може повністю заповнитися слизом і клітина відмирає. У деяких випадках слиз може проходити через первинну стінку клітини на поверхню. У синтезі та секреції слизу основну участь бере апарат Гольджі.
Слиз, що виділяється рослинними клітинами, виконує різні функції. Так, слиз кореневого чохлика служить як мастило, що полегшує зростання кінчика кореня в грунті. Слизові залозки комахоїдних рослин (росянка) виділяють ловчий слиз, до якого приклеюються комахи. Слиз, що виділяється зовнішніми клітинами насіннєвої шкірки (льон, айва, подорожники), закріплює насіння на поверхні ґрунту та захищає проросток від висихання. Слиз забарвлюється реактивом метиленовий синійу синій колір.
Виділення камедей зазвичай відбувається при пораненні рослин. Наприклад, камедетечення з поранених ділянок стовбурів та гілок часто спостерігається у вишні та сливи. Вишневий клей є застиглою камедь. Камедь виконує захисну функціюзакриваючи рану з поверхні. Утворюються камеді в основному у деревних рослин із сімейств бобових (акації, трагакантові астрагали) та розоцвітих підродини сливових (вишня, зливу, абрикос). Камеді та слизу використовуються в медицині.
Клітинна стінка є продуктом життєдіяльності протопласту. Полісахариди матриксу, глікопротеїд стінки, лігнін та слизу утворюються в апараті Гольджі. Синтез целюлози, освіта та орієнтація мікрофібрил здійснюються плазмалемою. Велика рольв орієнтації мікрофібрил належить мікротрубочкам, які розташовуються мікрофібрил, що паралельно відкладаються, поблизу плазмалеми. Якщо мікротрубочки зруйнувати, утворюються лише ізодіаметричні клітини.
Утворення клітинної стінки починається під час поділу клітини. У площині поділу утворюється клітинна платівка, єдиний шар, загальний двох дочірніх клітин. Вона складається з пектинових речовин, що мають напіврідку консистенцію; целюлоза відсутня. У дорослій клітині клітинна платівка зберігається, але зазнає змін, тому її називають серединний, або міжклітинною пластинкою (міжклітинною речовиною)(Рис. 2.16). Середня платівка зазвичай дуже тонка і майже невиразна.
Відразу після утворення клітинної платівки протопласти дочірніх клітин починають відкладати власну клітинну стінку. Вона відкладається зсередини як на поверхні клітинної пластинки, так і на поверхні інших клітинних стінок, що раніше належали материнській клітині. Після поділу клітина вступає у фазу росту розтягуванням, який зумовлений інтенсивним осмотичним поглинанням клітиною води, пов'язаним із утворенням та зростанням центральної вакуолі. Тургорний тиск починає розтягувати стіну, але вона не рветься завдяки тому, що в неї постійно відкладаються нові порції мікрофібрил та речовин матриксу. Відкладення нових порцій матеріалу відбувається рівномірно по всій поверхні протопласту, тому товщина стінки клітин не зменшується.
Стінки клітин, що діляться і ростуть, називають первинними. Вони містять багато (60-90%) води. У сухій речовині переважають матричні полісахариди (60-70%), вміст целюлози не перевищує 30%, відсутня лігнін. Товщина первинної стінки дуже невелика (01-05 мкм).
Для багатьох клітин відкладення клітинної стінки припиняється одночасно із припиненням росту клітини. Такі клітини оточені тонкою первинною стінкою до кінця життя ( Рис. 2.16).
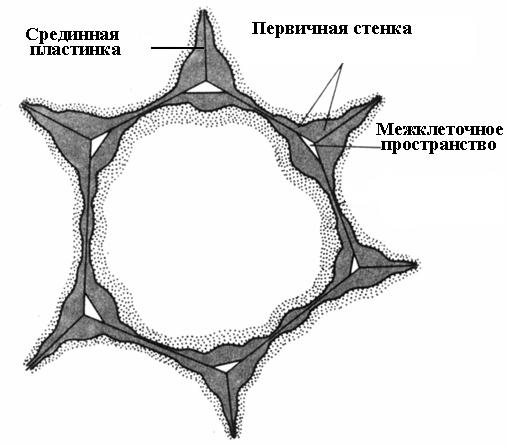
Рис. 2.16. Паренхімна клітка з первинною стінкою.
В інших клітин відкладення стінки продовжується і після досягнення клітиною остаточного розміру. При цьому товщина стінки збільшується, а об'єм, який займає порожнина клітини, скорочується. Такий процес має назву вторинного потовщеннястінки, а саму стінку називають вторинної(Рис. 2.17).
Вторинна стінка може розглядатися як додаткова, що виконує головним чином механічну опорну функцію. Саме вторинна стінка є відповідальною за властивості деревини, текстильного волокна, паперу. Вторинна стінка містить значно менше води, ніж первинна; в ній переважають мікрофібрили целюлози (40-50% від маси сухої речовини), які розташовуються паралельно один до одного. З полісахаридів матриксу характерні геміцелюлози (20-30%), пектинових речовин дуже мало. Вторинні клітинні стінки, як правило, зазнають здерев'янення. У неодревесневших вторинних стінках (луб'яні волокна льону, волоски бавовнику) вміст целюлози може досягати 95%. Великий змісті строго впорядкована орієнтація мікрофібрил визначають високі механічні властивостівторинних стін. Часто клітини, що мають вторинну одревесневу клітинну стінку, після завершення вторинного потовщення відмирають.
Середня платівка склеює сусідні клітини. Якщо її розчинити, стінки клітин втрачають зв'язок один з одним і роз'єднуються. Цей процес називається мацерація. Досить звичайна природна мацерація, при якій пектинові речовини серединної платівки переводяться в розчинний стан за допомогою ферменту пектинази і потім вимиваються водою (перестиглі плоди груші, дині, персика, банана). Часто спостерігається часткова мацерація, при якій серединна пластинка розчиняється не по всій поверхні, а лише у кутах клітин. Внаслідок тургорного тиску сусідні клітини у цих місцях округляються, внаслідок чого утворюються міжклітинні(Рис. 2.16). Міжклітини утворюють єдину розгалужену мережу, яка заповнюється парами води та газами. Таким чином, міжклітини покращують газообмін клітин.
Характерна рисавторинної стінки – її нерівномірне відкладення поверх первинної стінки, внаслідок чого у вторинній стінці залишаються непотовщені ділянки – пори. Якщо вторинна стінка не досягає великої товщини, пори виглядають як дрібні заглиблення. У клітин із потужною вторинною стінкою пори у розрізі мають вигляд радіальних каналів, що йдуть від порожнини клітини до первинної стінки. За формою порового каналу розрізняють пори двох типів – простіі про каймлені(Рис. 2.17).
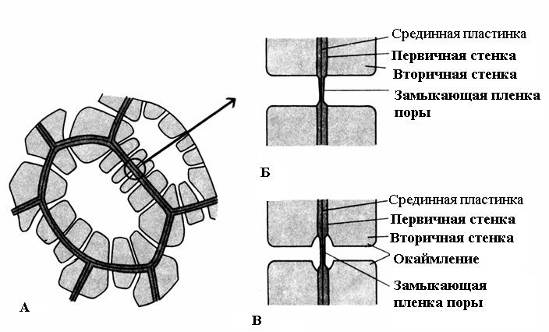
Рис. 2.17. Типи пір: А – клітини з вторинними стінками та численними простими порами; Б - пара простих пір; В – пара облямованих пір.
У простого часудіаметр порового каналу на всій довжині однаковий і має форму вузького циліндра. Прості пори характерні для паренхімних клітин, луб'яних та деревних волокон.
Пори у двох суміжних клітинах, як правило, виникають одна проти одної. Ці загальні пори мають вигляд одного каналу, розділеного тонкою перегородкою із серединної платівки та первинної стінки. Така сукупність двох пор суміжних стін сусідніх клітин носить назву пари пірта функціонує як одне ціле. Розділяючий їхній канал ділянка стінки називається замикаючої плівкоюпори, або поровий мембраною. У живих клітинах замикаюча плівка пори пронизана численними плазмодесмами(Рис. 2.18).
Плазмодесмипритаманні лише рослинним клітинам. Вони є тяжі цитоплазми, що перетинають стінку суміжних клітин. Число плазмодесм в одній клітці дуже велике – від кількох сотень до десятків тисяч, зазвичай плазмодесми зібрані до груп. Діаметр плазмодесменного каналу становить 30-60 нм. Його стінки вистелені плазмалемою, безперервною з плазмалемою суміжних клітин. У центрі плазмодесми проходить мембранний циліндр. центральний стриженьплазмодесми, безперервний з мембранами елементів ендоплазматичної мережі обох клітин Між центральним стрижнем та плазмалемою в каналі знаходиться гіалоплазма, безперервна з гіалоплазмою суміжних клітин.
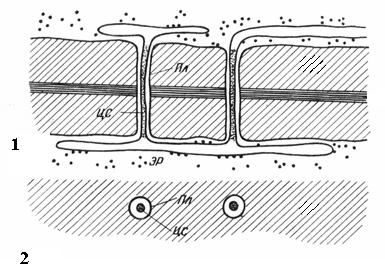
Рис. 2.18. Плазмодесми під електронним мікроскопом (схема): 1 - на поздовжньому зрізі; 2 – на поперечному зрізі; Пл- Плазмалема; ЦС– центральний стрижень плазмодесми; ЕР- Елемент ендоплазматичного ретикулуму.
Таким чином, протопласти клітин не повністю ізольовані один від одного, а повідомляються каналами плазмодесм. Ними відбувається міжклітинний транспорт іонів і дрібних молекул, і навіть передаються гормональні стимули. За допомогою плазмодесм протопласти клітин у рослинному організміутворюють єдине ціле, зване симпластом, а транспорт речовин через плазмодесми отримав назву симпластичногона відміну від апопластичноготранспорту по клітинних стінках та міжклітинниках.
У облямованих пір(Рис. 2.17)канал різко звужується в процесі відкладення клітинної стінки, тому внутрішній отвір пори, що виходить у порожнину клітини, набагато вже, ніж зовнішнє, що упирається в первинну стінку. Облямовані пори характерні для клітин водопровідних елементів деревини, що рано відмирають. У них поровий канал у напрямку замикаючої плівки воронковидно розширюється, а вторинна стінка нависає у вигляді валика над розширеною частиною каналу, утворюючи камеру пори. Назва облямованої пори походить від того, що при розгляді з поверхні внутрішній отвір має вигляд маленького кола або вузької щілини, тоді як зовнішній отвір як би облямовує внутрішній у вигляді кола більшого діаметра або ширшої щілини.
Пори полегшують транспорт у
Лекція: Особливості рослинної клітини
Сумарний хімічний склад майже однаковий.
Подібність із тваринною клітиною:наявність мембрани, цитозолю, ядра, та специфічних органел.
Однак між цими типами еукаріотів виділяються 2 відмінності в будові: наявність специфічних органоїдів та метаболізм.
У рослинній клітині є центральна вакуоля. Вакуолі рослинних клітин поглинають надлишок води, набухають, що призводить до розтягування клітин, вакуоля займає більшу частину об'єму рослинної клітини, і тому в раст.клетки залишаються лише тонкі тяжі цитоплазми, що проходять через усю клітину. Для растіт.клітини це служить економічним шляхом збільшення своїх розмірів. При цьому витрачається менше енергії, ніж у разі заповнення такого ж об'єму багатою на білки цитоплазмою.
У молодих рослинних клітин може бути кілька вакуолей, які зі зростанням клітини зливаються один з одним і утворюють одну або декілька великих вакуолей, які займають до 90% обсягу всієї клітини. При цьому органоїди відтісняються до периферії клітини і ядро теж. Мембрана вакуолі – тонопласт. Порожнина вакуолі заповнена клітинним соком, що є водним розчином неорганічних солей, цукрів, органічних кислот та їх солей, білки, низькомолекулярних сполук.
Головна функція центральної вакуолі- Підтримка тургорного тиску. Воно визначається розчиненими у клітинному соку вакуолі молекулами, що підтримують осмотичну концентрацію. Концентрація сприяє тому, що вакуоль функціонує як структура, що підтримує внутрішній тиск клітини, надаючи їй міцність і напруженість. У тонопласті є АТФ-залежний протонний насос, що забезпечує транспорт цукрів, а також там знаходяться іонні канали, які забезпечують екскрецію (виведення) з вакуолі різних метаболізмів. Концентрація протонів клітинному соку відповідає кислому середовищу 2-5 рН. Центральна вакуоль забезпечує накопичення запасних поживних речовин – цукрів та білків. Надходження цих речовин пов'язане із здатністю мембран взаємодіяти з тонопластом. Усередині вакуолі клітинного соку можуть здійснюватися і перетравлення, т.к. містяться гідролітичні ферменти, при цьому тонопласт формує секреторні бульбашки, вони відщеплюються від вакуолі, взаємодіють з екзоцетозними бульбашками, забезпечуючи перетравлення компонентів, що містяться в них. Вони виконують і реакції аутофатичного циклу, беручи участь у гідролізі дефектоного наповнення.
Клітинна стінка рослин.
Формується за участю плазмалеми. Є позаклітинним багатошаровим утворенням, що захищає поверхню клітини, будучи, по суті, його зовнішнім скелетом. Складається із двох структур: каркаса (з ниток фібрили целюлози) та матриксу гелеподібного. До складу матриксу входять полісахариди: геміцелюлози та пектинові речовини. Геміцелюлози являють собою полімерні ланцюги, що гілкуються, що складаються з різних гексоз (глюкоза, маноза, галактоза), можуть бути і пентози (ксилоза, арабінозу) і уронові кислоти (глюкуронова і галактуронова). Ці компоненти геміцелюлоз поєднуються між собою у різних кількісних відносинах, утворюють різноманітні комбінації. Але ланцюги геміцелюлозних молекул ніколи не кристалізуються, і вони не фіксуються (не виявляються) фібрили. Міститься висока кількість води.
Пектини. Гетерогенна група, в яку входять розгалужені, сильно гідротовані полімери, які несуть негативні зарядичерез наявність безлічі залишків галактуронової кислоти.
Завдяки цим компонентам матрикс є м'якою пластичною масою, яка є основою (каркасом).
Окремі полімери целюлози упаковані у мікрофібрили за допомогою водневих зв'язків. Надає жорсткості та міцності. Між ними існують зв'язки, які перетворюють клітинну стінку на моноліт. Такі пов'язані мікрофібрили целюлози оточені ще пектинами. Вони можуть взаємодіяти із кальцієм, кремнієм. Що надає жорсткості. За рахунок гелеподібної структури матрикс забезпечує дифузне пропускання води та невеликих молекул.
Клітинні стінки поділяються на 3 види: первинні, вторинні, третинні.
Первинна клітина складається на 90% вуглеводів. При розподілі клітин, в екваторіальній площині клітин з'являється пучок мікротрубочок, розташований між хромосомами, що розходяться (фрагмопласт). Серед мікротрубочок розташовується багато дрібних бульбашок вакуолярної системи, КГ, у центральній частині фрагмопласти починають зливатися один з одним. Утворюючи сплощений диск чи серединну платівку. Складається з полісахариду – калези. Більш еластичний порівняно з целюлозою (не міститься всередині бульбашок КГ, утворюється на плазмалемі). У бульбашках КГ містяться компоненти, необхідні для побудови мембран двох дочірніх клітин. Процес злиття дрібних вакуолей походить від центру клітини до периферії і продовжується до тих пір, поки мембранні бульбашки зливаючись не зіллються з поверхнею бічної поверхнею клітини.
Зростаюча первинна клітинна стінка складається вже з 3 шарів (серединна пластинка, що складається з аморфного матриксу, і 2 периферичних, що містять геміцелюлозу та фібрили). Первинна оболонка утворюється за рахунок виділення геміцелюлози та фібрил целюлози двома новими клітинними структурами. Збільшення товщини міжклітинної стінки відбуватиметься за рахунок активності дочірніх клітин, яка кожна зі свого боку виділятиме речовини клітинної оболонки, що потовщується шляхом підшаровування нових і нових пластів. На плазматичній мембрані синтезуються та полімеризуються целюлозні фібрили, що призводить до утворення вторинної клітинної стінки (надає клітині її остаточної форми). Як тільки потовщення стінки завершується відбувається її модифікація лігніном, гідрофобний полімер, синтезується за рахунок окисної полімеризації до трьох залишків ароматичних спиртів і служить основним компонентом деревини (20-35% деревини - лігнін). Завдяки своєму гідрофобному характеру лігнін створює гідроізоляцію в стінах і служить додатковим зміцнюючим матеріалом. Клітинна стінка дерев'яна, або опробковевает (за рахунок кутина і суберину). У клітинах епідерми лежить на поверхні клітинних оболонок виділяється віск.
У грибів хітин (N-ацетилглюкозамін).
Пластиди.
Загальна властивість всіх пластид:
Оточені двома мембранами, які тісно стуляються вздовж усієї поверхні органоїдів.
Внутрішній вміст – строма. У ньому мембранні диски, що утворилися в результаті вп'ячування і відбрунькування ділянок внутрішньої мембрани. Ці диски називаються тилакоїди, вони мають здатність складатися в стопки (грани). Вони взаємодіють із четвертим типом мембран – ламелою.
Пластиди мають власний геном, кожна пластида має множинні копії кільцевої ДНК, що містить близько 100 генів. Геном кодує білки, необхідних виконання спеціалізованих функцій. Тут же знаходяться білки РНК, що беруть участь у транскрипції та трансляції. Вони відбуваються в пластидах, але більшість інших білків залежить від ядерного геному. Ці білки синтезуються в цитоплазмі та повинні імпортуватися всередину пластид.
Пластиди пов'язані з іншими органоїдами везикулярними зв'язками.
Пластиди є рядом взаємних перетворень.
Пропластида -> лейкопласт -> хлоропласт -> хромопласт.
Лейкопласти можуть перетворюватися на:амілопласти, елайопласти, алейронові зерна та хромопласти.
Хлоропласт.
Мають подовжену форму, розмір 2-4мкм, а довжина може бути 10мкм. Хроматофори (величезні хлоропласти).
Внутрішній простір хлоропласту – люмен. 2 важливі явища: фотосинтез, синтез енергії. Характерним для мембрани тилакоїда є наявність хлорофілу, він здатний поглинати кванти сонячного світла та перетворювати енергію сонця на хімічну енергію. Поглинання світла з певною довжиною хвилі призводить до зміни хлорофілу. Він перетворюється на збуджений стан. І енергія активованого хлорофілу, що звільняється, через ряд проміжних етапів передається на ланцюг переносу електронів, що призводить до синтезу АТФ і відновлення переносника, яким є 2 мононуклеотиди (НАД і ФАД). Енергія витрачається на синтез діоксиду вуглецю та синтез цукрів.
Фотосинтез: світлова та темнова фаза.
Світлова фаза протікає тільки на світлі пов'язана з поглинання світла пігментом і проведенням хімічної реакції - реакція Хілла.
У темновій фазі відбувається фіксація та відновлення діоксиду вуглецю (з атмосфери), що призводить до синтезу вуглецю, АМК та інших речовин. В результаті світлової фази відбувається процес фосфорилування, синтез АДФ та АТФ. А також відновлення переносника, що відбувається при гідролізі та фотолізі води.
У світловій фазі фотосинтезу енергія сонячного світла збуджує електрони хлорофілу. Вони переносяться по компонентах окисного ланцюга в тилакоїдній мембрані, подібно до того, як електрони транспортуються дихальним ланцюгом в мітохондріях. У хлоропластах протони перекачуються з строми (рН = 8) в люмен (рН = 5) створюючи градієнт 3 одиниці рН. Цей протонний градієнт створює на протонній мембрані протон-рушійну силу 200 мВт, але вона майже цілком зумовлена градієнтом рН. У порівнянні з дихальним ланцюгом мітохондрій електрони рухаються в іншому напрямку. Електрони беруться від води, при її фотолізі та передаються на переносник із витратами енергії. Т.о. у світловій фазі ми маємо: синтезовані молекули АТФта відновлений переносник. Обидва продукти використовуються у темновій стадії.
У темновій стадії за рахунок відновленого НАД та АТФ відбувається утворення вуглеводів. Цей процес багатоступінчастий. У ньому бере участь велика кількість ферментів. Цикл Кальвіна. Збільшення числа хлоропластів та утворення інших форм пластид походить зі структур-попередників (пропластид).
Пропластиди дрібні двомембранні бульбашки, без рис. Вони відрізняються від вакуолей цитоплазми щільнішим вмістом, наявністю двох мембран. Пропластиди розташовуються в тканинах рослин, що діляться. Збільшується число пропластид шляхом поділу та брунькування. При нормальному освітленні пропластиди перетворюються на хлоропласти (збільшуються у розмірах, відбувається утворення поздовжньо розташованих мембранних складок. Одні формують ламели, інші – тилакоїди). Якщо вони розвиваються в темряві, то спочатку відбувається збільшення об'єму пластид, але система внутрішніх мембран не утворюється, утворюється маса дрібних бульбашок, які накопичуються в окремі зони, що формують складні ґратчасті структури – проламелярні тіла (етіопласти). У їх мембранах міститься протохлорофіл (жовтого кольору), під впливом світла їх утворюється хлоропласти і протохлорофіл може перетворитися на справжній хлорофіл (синтезується мембрани, компоненти ланцюга переносу).
Лейкопласти.Відрізняються від хлоропластів. Вони знаходяться в клітинах запасних тканин. Через них невизначену морфологію їх важко відрізнити від пропластиду. Вони видно ламеллами, але здатні до утворення нормальних тилакоїдних структур. У темряві вони накопичують різноманітні запасні речовини. Якщо вуглевод - то лейкопласти формуються в амілопласти (там можуть бути білки у формі алейронових зерен). А якщо жири – елайопласти.
Процес знебарвлення та зміни хлоропластів ми спостерігає при розвитку пелюсток та дозріванні плодів. У пластидах накопичується доп.мембрани, руйнується хлорофіл і крохмаль. При руйнуванні ламелл виділяються ліпідні краплі, у яких добре розчиняються пігменти (каратиноїди) та проявляється зміна фарбування. Хромопласти є дегенеруючою формою пластид, що пов'язано з розпадом ліпопротеїдних комплексів.
Геном пластид.
Власна генетична система забезпечує синтез білків, які локалізуються усередині самих пластид (білки тилакоїдної мембрани). Мають довжину до 60 мкм. 1.3 * 10 8 Так. Тривалість циклу та швидкість реплікації не збігається. ДНК немає фіксуючих структур. Всі властивості кільцевої молекули хлоропластів близькі до параметрів прокаріотів. Подібність ДНК хлоропластів проявляється у тому, що основні регуляторні процеси транскрипції вони однакові. Синтез усіх видів РНК. ДНК хлоропластів кодує РНК рибосом. Константи сегментації рибосом 70S. Вони містять 17S та 23S. Чутливі до антибіотика (хлорамфенікол).
Лекція: Міжклітинні контакти
Симбіотична теорія.
Комінцин та Мерешковський.
На користь цієї теорії говорить дивовижна подібність у будові хлоропластів та синьо-зелених водоростей. Подібність до основних їх функцій, практично однакові здібності до фотосинтетичних процесів. Внаслідок симбіозу різних прокаріотичних клітин виникли ядерні (еукаріотичні) клітини. Відповідно до цієї гіпотизи еукаріотична клітина у своєму розвитку пройшла кілька етапів симбіозу з іншими клітинами.
На першій стадії клітини типу анаеробних гетеротрофних прокаріотів стали господарями для аеробних бактерій, що призвело до утворення аеробних прокаріотів (мітохондрій, усередині цих прокаріотичних організмів). Паралельно з цим процесом у клітині-господарі прокаріотичний ядерний апарат (генофор) формується в відокремлене від цитоплазми ядро. Так могли з'явитися гетеротрофні еукаріотичні клітини. Така клітина входить у повторний симбіоз. Наприклад, зі спірохетами чи джгутикоподібними бактеріями. Включення цих симбіонтів призвело до виникнення джгутиків та вій у гетеротрофних еукаріотичних клітин. І внаслідок додаткового поглинання. Це призвело до виникнення тварин еукаріотичних клітин, що мають джгутики. Такий організм міг поглинути і синьо-зелені водорості, і це призвело до утворення пластид та утворення рослинної клітини. Гіпотеза ендосімбіотичного походження внутрішньомембранних органоїдів не викликає сумніву. Відомі численні факти справжнього ендосимбіозу синьо-зелених водоростей і клітин нижчих рослин і найпростіших, де симбіонти чудово функціонують, постачають клітину-господаря продуктами фотосинтезу. Наприклад. Інфузорія-туфелька, в ній живе водорість хлорела. Хлоропласти можуть відбиратися деякими клітинами та використовуватися як ендосімбіонти. Коловратки харчуються водоростями. Хлоропласти виявляються усередині клітин травних залоз та продовжують виконувати свої функції.
Особливості функцій рослинних клітин. Спосіб синтезу органічних речовин – автотрофне харчування.
Клітини рослин за рахунок наявності клітинної стінки не дозволяють рослинній клітині змінювати свою форму та переміщатися. І клітини рослин компенсують відсутність руху тим, що синтезують собі їжу, тобто. є автотроф. Рослини ростуть протягом усього життя, меристеми постійно діляться, утворюють нові клітини, всі рослини відносяться до модулярних організмів. Їхня остаточна життєва форма залежить від навколишнього середовища. Клітини рослин мають особливості і щодо протікання мітозу:
Положення нової клітинної стінки. Визначається задовго до утворення веретена, при цьому саме веретено не бере участі у розташуванні цієї клітини. Ще до мітозу G 2 фазі клітинного циклу утворюється специфічна структура в рослинній клітині, яка називається препрофазне кільце. Формується воно з мікротрубочок кортексу (цитоскелета) у вигляді широкої смужки, що оточує клітину. Ядро залишається інтактним (цілісним). В області цього кільця збираються актинові філаменти, які забезпечують його звуження доти, доки воно не перетвориться на добре оформлений щільний пучок мікротрубочок навколо ядра. Структура, що вийшла, нагадує колесо, в якому обід і спиці складаються з мікротрубочок і актину, а ядро утворює втулку. Структура кільця збагачена елементами ЕПР та КГ. На ранній стадії мітозу мікротрубочки препрофазного кільця деполіміризуються та руйнуються. І в телофазі нова клітинна стінка утворюється точно в площині, яка була позначена препрофазним кільцем.
Ядро в рослинній клітині рухається в центр клітини, де відбуватиметься поділ. Від ядра на всі напрямки поширюються вузькі шари (тяжі) цитоплазми. У міру настання мітозу шари зливаються між собою, утворюючи площину там, де раніше було препрофазне кільце, і ця площина називається фрагмосома.
У рослин полюси мітотичного веретена не містять центріолей і відрізняються більш дифузним характером. Тобто вони ніби розмиті. На початку з мікротрубочок формується профазне веретено. З нього після руйнування ядерної оболонкиутворюється вже справжнє веретено, причому при утворенні цього справжнього мітотичного веретена в клітині взагалі не залишається інших цитоплазматичних мікротрубочок. Що керує цим веретеном, чим утворюється? Хромосоми. Спочатку поблизу хромосом відбувається полімеризація мікротрубочок, які орієнтуються випадково. Потім моторні білки (дінеїн) сортують мікротрубочки, їх позитивні кінці направляються в хромосоми, що дозволяє приєднуватися до кінетохорів хромосом, а з негативними кінцями мікротрубочок взаємодіють інші. Внаслідок полімеризації, сортування, прикріплення утворюється біполярне веретено. Після розподілу ядра настає розподіл цитоплазми. Що збігається з моментом закладання нової клітинної перегородки, становище якої визначається препрофазним кільцем. Зв'язок між орієнтацією веретена і новою перегородкою немає тісного характеру, як і клітинах тварин.
Гольджі рослинних клітин комплексу. Функції: глікозилювання (первинне), сортинг (поділ), синтез полісахаридів клітинної стінки (крім целюлози та колізи), доставка на мембрану ферментних комплексів (гексагональних розеток, синтез целюлози), спрямований транспорт везикул у вакуолі.
Локалізовано КГ у рослинних клітинах ближче до плазми, далі від ядра, і розосереджено у вигляді численних стосів цистерн. Усі елементи комплексу пов'язані актиновими нитками, що утворюють мережу, що забезпечує внутрішньоклітинний транспорт. Актинові структури забезпечують рухливість (разом із міозином). У клітинах рослин є особливий міозин - міозин 8. Цей міозин 8 не утворює філаментів і накопичується в первинних первинних стінках, у везикулах КГ містяться полісахариди і структурні білки, що формують вторинну клітинну стінку.
Міжклітинні контакти.
Контактна функція мембран забезпечує контакт клітин між собою та з іншими. Усі клітини у тканинах перебувають у контакті з позаклітинним матриксом. Позаклітинний матрикс бере участь у підтримці цілісності тканини і утворює впорядкований кістяк, усередині якого клітини і переміщаються та взаємодіють один з одним. Взаємодія клітин здійснюється за рахунок дотичних ділянок, які називаються міжклітинними контактами.
Їх класифікацій є кілька, по-різному їх структурують. Класифікація Альбертса та Грея (1986). Відповідно до неї: контакти можна розбити на 3 групи.
Механічні (= Адгезійні). Пов'язані зі здатністю клітин злипатися один з одним, завдяки наявності в мембрані клітин глікопротеїнів позаклітинного матриксу. При адгезії між плазматичними мембранами залишається щілина (20нм) заповнена глікокаліксом. Безпосередньо за з'єднання клітин одна з одною відповідають спеціальні білки САМ (cam): фібронектин, кадгерини, селектини, інтегрини та інші білки. Деякі їх пов'язують клітини друг з одним з допомогою межмолекулярных взаємодій. Інші утворюють спеціальні міжклітинні сполуки. Взаємодія між цими білками може бути гомофільними, тобто. сусідні клітини у разі зв'язуються друг з одним з допомогою однорідних молекул. Або гетерофільні, коли в адгезії беруть участь різного родубілки на сусідніх клітинах. Бувають і складніші сполуки, коли міжклітинне зв'язування забезпечується з допомогою лінкерних проміжних молекул. Крім таких простих зв'язківіснує ряд спеціальних структурякі виконують конкретні функції (десмосоми). Вони взаємодіють із елементами цитосклета. Виділяють 3 різновиди десмосом: точкові, оперізуючі та напівдесмосоми.
Точкові з'єднують клітини одна з одною у вигляді кнопок. Відстань між мембранами контактуючих клітин варіює від 22 до 35 нм. Між клітинами за рахунок видозмін надмембранного комплексу формується волокнистий матрикс і в його центральній частині розташовується пластинка, що складається з білкових глобул. Вони представлені взаємодіючими інтегральними мембранними білками каддеринами та десмоглеїнами. Ця платівка взаємодіє з мембранами клітин системою поперечних тяжів. З боку цитоплазми формується друга пластинка з білка десмоплакіна, від якого відходить у поперечному напрямку вглиб фібрилярні структури, що відносяться до 10нм філаментів (кератинові волока). Звідси точкові десмосоми зустрічаються в епітелії, серцевих м'язах, ендотельних судин.
Оперізуючі десмосоми. Відстань між сусідніми клітинами 15-20 нм. Центральної платівки такої яскравої немає. Але є транс-мембранні глікопротеїни, які зчіплюються один з одним та забезпечують механічне з'єднання мембран. З боку цитоплазми накопичуються тонкі актинові філаменти (6-7нм). Вони лежать уздовж плазмалеми і йдуть по всьому периметру клітини. Функція: механічне зчеплення клітин одна з одною; при скороченні актинових філаментів може змінюватись форма клітини; може викликати зміну геометрії клітини; передають механічну напругу клітини. Викликане зміною об'єму клітини.
Напівдесмосоми. З'єднання клітин з міжклітинним матриксом (Наприклад, з базальною мембраною). Функціональна роль– механічна, що дозволяє витримувати епітеліальним пластам великі механічні дії. Десмосоми – ділянки клітин, але де вони статичні освіти. Вони динамічні. Чи здатні зникати, знову з'являтися. Знищуються шляхом фагоцентарної реакції.
Замикаючі (щільні контакти). Вони характерні для одношарових епітеліїв. Зона, де зовнішні шари двох плазматичних мембран максимально зближені. Тому при перегляді в електронний мікроскоп видно тришарову структуру. 2 зовнішні шари обох мембран зливаються в один товщиною 2-3нм. Не по всій площі щільного контакту, а є рядом точкових дотиків інтегральних білків мембран. У зоні щільних контактів виявлено понад 24 види білків. Виступаючи з мембран біліпідного шару, такі білки перетинаються, утворюють мережу (решітку). З боку щільного контакту в самій цитоплазмі розташовуються багато чисельних фібрил (7нм), які розташовуються паралельно поверхні плазмалеми. Щільні контакти характерні для залізистих та кишкових епітелій. Вони з'єднують клітини одна з одною. Крім того, це молекулярні структури, Регулюючі параклеточный транспорт, тобто. транспорт речовин у просторі між клітинами. Фізичні бар'єри транспорту іонів мають різну природу. Іони проходять миттєво, інші компоненти для проходження через щільний контакт витрачають більше часу (навіть годинник).
Бар'єр проникності в щільному контакті створюється за рахунок шарів, що несуть заряди, що утворюють сітчасту структуру фібрилярних ниткоподібних елементів. Іони можуть проходити через такі пори, їх розмір невеликий. Однак для проходження розчинних компонентів повинна порушуватися цілісність цих ниток. Розчинні елементи розривають ці нитки, потім вони з'єднуються після проходження молекул. І так молекула просувається через бар'єр контакту. У структурному та функціональному відношенні щільні контакти ділять плазмолемму на 2 ділянки (апікальну та базальну) і щільний контакт виконує роль перегородки, за рахунок якої підтримується молекулярний склад у межах.
Провідні контакти. По-різному у тварин та рослин.
У тварин клітинах 2 типи: щілинні та хімічний синтез. Через щілинні контакти молекули можуть переходити з однієї клітини до іншої. А у хімічному синтезі клітини не мають безпосереднього зв'язку. Щілинні контакти вважають комунікаційними (сполученими) сполуками клітин. Це структура, яка бере участь у прямій передачі хімічних речовин. Характерним для цього контактом є зближення плазматичних мембран двох сусідніх клітин на відстань 2-3 нм. Сам щілинний контакт має розмір 0,5-5мкм. Складається з 2 половин, розмір кожної половинки 7-8нм, розташовуються навколо каналу шириною 1-2 нм. І ці дві половинки називаються конексони (напівканали). Вони складаються з шести субодиниць білка коннектину. Молекулярна маса білка близько 30 тисяч. Об'єднуючись один з одним білки формують циліндричну структуру і двох клітин величини, число, розмір, розташування конексонів суворо симетричний. Щілинні контакти допускають вільну дифузію молекул, масою 1200 Дальтон, виключають молекули маса якої досягає 2000 Дальтон. Проникність щілинних контактів регулюється відкриттям та закриттям воріт каналу. Цей процес має назву гейтинг (воротний механізм). Він контролюється зміною внутрішньоклітинного рН, потоком іонів кальцію або прямим фосфорилуванням субодиниць білка коннексину. Функції щілинних контактів: проведення органічних та неорганічних речовин. (Наприклад, у м'язових клітинах міокардів).
Різновидом провідних контактів рослин є плазмодесми. Це тонкі трубчасті цитоплазматичні канали, що з'єднують дві сусідні клітини. Діаметр цих каналів варіює від 20 до 40 нм. Особливість цих каналів: плазматична мембранаоднієї клітини перетворюється на плазмалемму інший клітини. По осі цього каналу з 1 клітини до іншої тягнеться циліндрична трубочка десмотула. Просвіток її повідомляється з ЕПР клітин. Простір між десмотубулою та мембраною плазмодесми заповнено цитозолем. Плазмалема, що обмежує ці клітини, плавно переходить у плазмодесму і мембрану інших клітин. В результаті відбувається поєднання гіалоплазми сусідніх клітин в єдину систему (симпласт). І формально поділу немає (однієї клітини від іншої). Вільний транспорт через плазмодесму обмежується молекулярною масою 800 Дальтон. Утворюється плазмодесма ще на етапі первинного формування серединної платівки з елементів ЕПР. При цьому у клітини, що тільки що розділилася, число плазмодесм досягає до 1000 на клітину. При старінні їх кількість скорочується, але збільшується товщина. Функціональна роль: забезпечення міжклітинної циркуляції розчинів (іони, цукри, поживні речовини). Негативна сторона – захворіти легше.
ВАКУОЛІ РОСЛИННИХ КЛІТИН
Органоїди загального значення, що мають одномембранний принцип будови та зустрічаються тількиу клітинах рослин.
Розмір та кількість: визначаються віком клітини. У молодих клітинах вакуолі виникають з дрібних бульбашок, що відщепилися від ЕПС. У міру зростання та диференціювання клітини дрібні вакуолі зливаються один з одним і утворюють одну або кілька великих вакуолей, що займають до 80% обсягу всієї клітини. В результаті цитоплазма з ядром та органоїдами відтісняється на периферію.
Структура: центральні вакуолі відокремлені від цитоплазми одинарною мембраною тонопластом, Який подібний за товщиною з плазмолемою. Порожнина вакуолі заповнена клітинним соком. До складу клітинного соку входять неорганічні солі, цукру, органічні кислоти та їх солі, інші низькомолекулярні речовини, а також деякі високомолекулярні сполуки (наприклад, білки).
Функції: 1) осморегуляція: завдяки напівпроникності тонопласту та плазмолеми зберігається відповідна молекулярна концентрація клітинного соку, тобто. вакуоль функціонує як осмометр;
2) екскреторна: через тонопласт можуть видалятися всі водорозчинні продукти обміну (алкалоїди – нікотин, кофеїн; поліфеноли);
3) запасаюча: у клітинному соку накопичуються фосфати K + , Na + , Ca 2+ , солі органічних кислот (оксалати, цитрати та ін), цукру та білки.
РИБОСОМИ
Органоїд, що не має мембранної будови. Це єдиний органоїд загального значення, який присутній у клітинах як прокаріотів, так і еукаріотів. Рибосоми вперше були описані в 1955 м. Дж. Паладе (гранули Паладе), який довів, що вони є рибонуклеопротеїдними комплексами (РНП).
Хімічна природа: РНП= Р-РНК + білок.
На частку рибосом доводиться 85% РНКпредставлена в клітині.
Форма: рибосома має грибоподібну форму, оскільки складається з двох субодиниць: великийі малої, між ними розташовується функціональний центр рибосоми (ФЛР) , В якому під час біосинтезу білка (період трансляції) розташовується і-РНК своїми двома триплетами і працює ферментативний комплекс, що забезпечує складання білкової молекули з амінокислот.
розміри: 15 – 35 нм. Розмір повної рибосоми прокаріотичних клітин – 20х17х17 нм, еукаріотичних – 25х20х20 нм.
Місце освіти: Утворення субодиниць рибосом відбувається в ядерцях ядра. Складання субодиниць у цілісну рибосому здійснюється в цитоплазмі при досягненні концентрації іонів магнію (Mg 2+) 0.001М, якщо зазначена концентрація зменшується, відбувається дисоціація субодиниць. Коли концентрація Mg 2+ збільшується вдесятеро, досягаючи значення 0.01М, дві рибосоми взаємодіють один з одним, утворюючи димер.


